Subiectul și sarcinile imunologiei. Etape istorice în dezvoltarea imunologiei. Etape de formare a imunologiei Bariera protectoare a tractului gastrointestinal
Imunologia ca domeniu specific de cercetare a apărut din necesitatea practică de a combate bolile infecțioase. Ca direcție științifică separată, imunologia s-a format abia în a doua jumătate a secolului al XX-lea. Istoria imunologiei ca ramură aplicată a patologiei infecțioase și microbiologiei este mult mai lungă. Observațiile vechi de secole ale bolilor infecțioase au pus bazele imunologiei moderne: în ciuda răspândirii pe scară largă a ciumei (secolul al V-lea î.e.n.), nimeni nu s-a îmbolnăvit de două ori, cel puțin mortal, iar cei care și-au revenit au fost folosiți pentru a îngropa cadavrele.
Există dovezi că primele vaccinări împotriva variolei au fost efectuate în China cu o mie de ani înainte de nașterea lui Hristos. Inocularea conținutului de pustule de variolă la oameni sănătoși pentru a le proteja de forma acută a bolii s-a răspândit apoi în India, Asia Mică, Europa și Caucaz.
Inocularea a fost înlocuită cu metoda de vaccinare (din latinescul „vacca” - vacă), dezvoltată la sfârșitul secolului al XVIII-lea. doctor englez E. Jenner. El a atras atenția asupra faptului că asistentele care îngrijeau animalele bolnave se îmbolnăveau uneori într-o formă extrem de ușoară de variola bovină, dar nu s-au îmbolnăvit niciodată de variolă. O astfel de observație a oferit cercetătorului o oportunitate reală de a lupta împotriva bolii oamenilor. În 1796, la 30 de ani de la începutul cercetărilor sale, E. Jenner a decis să testeze metoda de vaccinare împotriva variolei bovine. Experimentul a avut succes și de atunci metoda de vaccinare conform lui E. Jenner a fost utilizată pe scară largă în întreaga lume.
Nașterea imunologiei infecțioase este asociată cu numele unui remarcabil om de știință francez Louis Pasteur. Primul pas către o căutare țintită a preparatelor de vaccin care creează imunitate stabilă la infecție a fost făcut după observarea lui Pasteur a patogenității agentului cauzal al holerei la pui. Din această observație, Pasteur a concluzionat că o cultură în vârstă, care și-a pierdut patogenitatea, rămâne capabilă să creeze rezistență la infecție. Acest lucru a determinat timp de multe decenii principiul creării materialului de vaccin - într-un fel sau altul (pentru fiecare agent patogen în felul său) pentru a obține o scădere a virulenței agentului patogen, menținându-și în același timp proprietățile imunogene.
Deși Pasteur a dezvoltat principiile vaccinării și le-a pus în practică cu succes, nu era conștient de factorii implicați în procesul de protecție împotriva infecțiilor. Primii care au făcut lumină asupra unuia dintre mecanismele de rezistență la infecție au fost Emil von Behringși Kitazato. Ei au demonstrat că serul de la șoareci imunizați anterior cu toxină tetanica, administrat animalelor intacte, le-a protejat de o doză letală de toxină. Factorul seric, o antitoxină, formată în urma imunizării, a fost primul anticorp specific descoperit. Lucrările acestor oameni de știință au marcat începutul studiului mecanismelor imunității umorale.
La originile cunoașterii problemelor imunității celulare a fost un biolog evoluționist rus Ilia Ilici Mechnikov. În 1883, el a făcut primul raport despre teoria fagocitară a imunității la un congres al medicilor și oamenilor de știință naturală de la Odesa. O persoană are celule mobile ameboide - macrofage, neutrofile. Ei „mănâncă” alimente de un fel special - microbi patogeni, funcția acestor celule este lupta împotriva agresiunii microbiene.
În paralel cu Mechnikov, un farmacolog german și-a dezvoltat teoria protecției imune împotriva infecțiilor Paul Erlich. Era conștient de faptul că în serul sanguin al animalelor infectate cu bacterii apar substanțe proteice care pot ucide microorganismele patogene. Aceste substanțe au fost ulterior denumite de el „anticorpi”. Proprietatea cea mai caracteristică a anticorpilor este specificitatea lor pronunțată. Formați ca agent de protecție împotriva unui microorganism, îl neutralizează și îl distrug doar pe acesta, rămânând indiferenți față de ceilalți.
Două teorii - fagocitară (celulară) și umorală - în perioada apariției lor au stat pe poziții antagonice. Școlile lui Mechnikov și Erlich s-au luptat pentru adevărul științific, fără a bănui că fiecare lovitură și fiecare paradă își apropiau adversarii. În 1908, ambii oameni de știință au primit simultan Premiul Nobel.
La sfârșitul anilor 1940 și începutul anilor 1950, prima perioadă de dezvoltare a imunologiei se apropia de sfârșit. A fost creat un întreg arsenal de vaccinuri împotriva celei mai largi game de boli infecțioase. Epidemiile de ciumă, holeră, variolă au încetat să distrugă sute de mii de oameni. Încă mai apar focare individuale, sporadice, ale acestor boli, dar acestea sunt doar cazuri foarte locale care nu au semnificație epidemiologică, cu atât mai puțin pandemică.
Orez. 1. Imunologi: E. Jenner, L. Pasteur, I.I. Mechnikov, P. Erlich.
O nouă etapă în dezvoltarea imunologiei este asociată în primul rând cu numele unui remarcabil om de știință australian. M.F. Burnet. El a fost cel care a determinat în mare măsură fața imunologiei moderne. Considerând imunitatea ca o reacție care vizează diferențierea tot ceea ce „al propriu” de tot ceea ce „străin”, el a pus problema semnificației mecanismelor imunitare în menținerea integrității genetice a organismului în perioada dezvoltării individuale (ontogenetice). Burnet a fost cel care a atras atenția asupra limfocitului ca principal participant la un răspuns imun specific, dându-i numele de „imunocit”. Burnet a fost cel care a prezis și englezul Peter Medawarși cehă Milan Hasek a confirmat experimental starea opusă reactivității imune – toleranță. Burnet a fost cel care a subliniat rolul special al timusului în formarea răspunsului imun. Și, în cele din urmă, Burnet a rămas în istoria imunologiei ca creator al teoriei selecției clonale a imunității. Formula unei astfel de teorii este simplă: o clonă de limfocite este capabilă să răspundă doar la un determinant specific, antigenic, specific.
De remarcat sunt părerile lui Burnet despre imunitate ca reacție a unui astfel de organism care distinge totul „propriu” de tot ceea ce „străin”. După ce Medawar a dovedit natura imunologică a respingerii unei grefe străine, după acumularea de fapte privind imunologia neoplasmelor maligne, a devenit evident că răspunsul imun se dezvoltă nu numai la antigenele microbiene, ci și atunci când există, deși nesemnificative, antigenice. diferențe dintre organism și acel material biologic (grefă, tumoră malignă) cu care se întâlnește.
Astăzi știm, dacă nu toate, atunci multe dintre mecanismele răspunsului imun. Cunoaștem baza genetică pentru o varietate surprinzător de mare de anticorpi și receptori de recunoaștere a antigenului. Știm ce tipuri de celule sunt responsabile pentru formele celulare și umorale ale răspunsului imun; mecanismele de creștere a reactivității și toleranței sunt în mare măsură înțelese; se cunosc multe despre procesele de recunoaștere a antigenelor; au fost identificați participanți moleculari ai relațiilor intercelulare (citokine); în imunologia evoluţionistă s-a format conceptul rolului imunităţii specifice în evoluţia progresivă a animalelor. Imunologia, ca ramură independentă a științei, a devenit la egalitate cu adevăratele discipline biologice: biologie moleculară, genetică, citologie, fiziologie și doctrina evoluționistă.
/ 62
Cel mai rau Cel mai bun

Imunologia a apărut ca parte a microbiologiei ca urmare a aplicării sale practice pentru tratamentul bolilor infecțioase, astfel încât imunologia infecțioasă sa dezvoltat în prima etapă.
De la începuturile sale, imunologia a interacționat strâns cu alte științe: genetică, fiziologie, biochimie și citologie. În ultimii 30 de ani, a devenit o știință biologică fundamentală vastă, independentă. Imunologia medicală rezolvă practic majoritatea problemelor de diagnostic și tratament al bolilor și în acest sens ocupă un loc central în medicină.
La originile imunologiei stau observațiile popoarelor antice. În Egipt și Grecia, se știa că oamenii nu s-au mai îmbolnăvit de ciumă și, prin urmare, cei care fuseseră bolnavi erau implicați în îngrijirea bolnavilor. Cu câteva secole în urmă, în Turcia, Orientul Mijlociu și China, puroiul din ulcerele uscate de variolă era frecat în piele sau mucoasele nasului pentru a preveni variola. O astfel de infecție a provocat de obicei o formă ușoară de variolă și a creat imunitate la reinfecție. Această metodă de prevenire a variolei se numește variolare. Cu toate acestea, mai târziu s-a dovedit că această metodă este departe de a fi sigură, deoarece uneori duce la variolă severă și moarte.
Din cele mai vechi timpuri, oamenii au știut că pacienții care au avut variola bovină nu se îmbolnăvesc de variola. Timp de 25 de ani, medicul englez E. Jenner a verificat aceste date cu numeroase studii și a ajuns la concluzia că infecția cu variola bovină previne boala variolei. În 1796, Jenner a altoit material din abcesul de variolă al unei femei infectate cu variola bovină într-un băiețel de opt ani. Câteva zile mai târziu, băiatul a făcut febră și au apărut abcese la locul de injectare a materialului infecțios. Apoi aceste fenomene au dispărut. După 6 săptămâni i s-a injectat material pustular de la un pacient cu variolă, dar băiatul nu s-a îmbolnăvit. Cu această experiență, Jenner a stabilit mai întâi posibilitatea de a preveni variola. Metoda a devenit larg răspândită în Europa, rezultând o scădere bruscă a incidenței variolei.
Metodele bazate pe dovezi pentru prevenirea bolilor infecțioase au fost dezvoltate de marele om de știință francez Louis Pasteur. În 1880, Pasteur a studiat holera la pui. Într-unul dintre experimentele sale, a folosit o cultură veche a agentului cauzal al holerei de pui, care a fost păstrată pentru o lungă perioadă de timp la o temperatură de 37 ° C, pentru a infecta puii. Unii dintre puii infectați au supraviețuit și după reinfectare. cu o cultură proaspătă, găinile nu au murit. Pasteur a raportat acest experiment Academiei de Științe din Paris și a sugerat că microbii slăbiți ar putea fi folosiți pentru a preveni bolile infecțioase. Culturile slăbite au fost numite vaccinuri (Vacca - vacă), iar metoda de prevenire - vaccinare. Ulterior, Pasteur a primit vaccinuri împotriva antraxului și a rabiei. Principiile de obținere a vaccinurilor dezvoltate de acest om de știință și metodele de aplicare a acestora au fost folosite cu succes de 100 de ani pentru prevenirea bolilor infecțioase. Cu toate acestea, cum este creată imunitatea nu se știe de mult timp.
Dezvoltarea imunologiei ca știință a fost mult facilitată de cercetările lui I. I. Mechnikov. Prin studii, I. I. Mechnikov a fost zoolog, a lucrat la Odesa, apoi în Italia și Franța, la Institutul Pasteur. În timp ce lucra în Italia, a experimentat cu larve de stele de mare, pe care le-a injectat cu spini de trandafiri. În același timp, el a observat că celulele mobile s-au acumulat în jurul vârfurilor, învăluindu-le și captându-le. I. I. Mechnikov a dezvoltat teoria fagocitară a imunității, conform căreia eliberarea organismului de microbi are loc cu ajutorul fagocitelor.
A doua direcție în dezvoltarea imunologiei a fost reprezentată de omul de știință german P. Ehrlich. El credea că principalul mecanism de protecție împotriva infecției sunt factorii umorali ai serului sanguin - anticorpi. Până la sfârșitul secolului al XIX-lea, a devenit clar că aceste două puncte de vedere nu se exclud, ci se completează reciproc. În 1908, I. I. Mechnikov și P. Ehrlich au primit Premiul Nobel pentru dezvoltarea doctrinei imunității.
Ultimele două decenii ale secolului al XIX-lea au fost marcate de descoperiri remarcabile în domeniul microbiologiei și imunologiei medicale. Serurile antitoxice pentru tetanos și difterie au fost obținute prin imunizarea iepurilor cu toxină difterice și tetanica. Așadar, pentru prima dată în practica medicală, a apărut un remediu eficient pentru tratamentul și prevenirea difteriei și tetanosului. În 1902, Behring a primit Premiul Nobel pentru această descoperire.
În 1885, Buchner și colegii de muncă au descoperit că microbii nu se înmulțesc în serul de sânge proaspăt, adică are proprietăți bacteriostatice și bactericide. Substanța conținută în ser, când a fost încălzită și păstrată timp îndelungat, a fost distrusă. Ehrlich a numit mai târziu această substanță complement.
Omul de știință belgian J. Borde a arătat că proprietățile bactericide ale serului sunt determinate nu numai de complement, ci și de anticorpi specifici.
În 1896, Gruber și Durham au descoperit că atunci când animalele sunt imunizate cu diverși microbi, în ser se formează anticorpi care provoacă aderența microbilor (aglutinarea). Aceste descoperiri au extins înțelegerea mecanismelor de protecție antibacteriană și au făcut posibilă aplicarea reacției de aglutinare în scopuri practice. Încă din 1895, Vidal folosea testul de aglutinare pentru a diagnostica febra tifoidă. Ceva mai târziu, au fost dezvoltate metode serologice pentru diagnosticarea tularemiei, brucelozei, sifilisului și a multor alte boli, care sunt utilizate pe scară largă în clinica bolilor infecțioase în prezent.
În 1897, Krause a descoperit că, pe lângă aglutinine, atunci când animalele sunt imunizate cu microbi, se formează și precipitine, care se combină nu numai cu celulele microbiene, ci și cu produsele metabolismului lor. Ca urmare, se formează complexe imune insolubile, care precipită.
În 1899, Ehrlich și Morgenrot au stabilit că eritrocitele adsorb anticorpi specifici pe suprafața lor și se lizează atunci când li se adaugă complement. Acest fapt a fost important pentru înțelegerea mecanismului reacției antigen-anticorp.
Începutul secolului al XX-lea a fost marcat de o descoperire care a transformat imunologia dintr-o știință empirică într-una fundamentală și a pus bazele dezvoltării imunologiei non-infecțioase. În 1902, omul de știință austriac K. Landsteiner a dezvoltat o metodă de conjugare a haptenelor cu purtători. Acest lucru a deschis posibilități fundamental noi pentru studierea structurii antigenice a substanțelor și a proceselor de sinteză a anticorpilor. Landsteiner a descoperit izoantigenele eritrocitelor umane din sistemul ABO și grupele sanguine. A devenit clar că există o eterogenitate în structura antigenică a diferitelor organisme (individualitate antigenică) și că imunitatea este un fenomen biologic care este direct legat de evoluție.
În 1902, oamenii de știință francezi Richet și Portier au descoperit fenomenul de anafilaxie, pe baza căruia a fost creată ulterior doctrina alergiilor.
În 1923, Gleny și Ramon au descoperit posibilitatea transformării exotoxinelor bacteriene sub influența formolului în substanțe netoxice - toxoizi cu proprietăți antigenice. Acest lucru a permis utilizarea toxoidelor ca preparate de vaccin.
Metodele de cercetare serologică sunt utilizate în altă direcție - pentru clasificarea bacteriilor. Folosind seruri antipneumococice, Griffith în 1928 a împărțit pneumococii în 4 tipuri, iar Lensfield, folosind antiseruri împotriva antigenelor specifice grupului, a clasificat toți streptococii în 17 grupe serologice. Multe tipuri de bacterii și viruși au fost deja clasificate în funcție de proprietățile lor antigenice.
O nouă etapă în dezvoltarea imunologiei a început în 1953 cu studiile oamenilor de știință britanici Billingham, Brent, Medavar și savantului ceh Hasek asupra reproducerii toleranței. Pe baza ideii prezentate în 1949 de Burnet și dezvoltată în continuare în ipoteza lui Jerne că abilitatea de a distinge între antigenele de sine și cele străine nu este înnăscută, ci se formează în perioadele embrionare și postnatale, Medawar și colegii săi au obținut toleranță la începutul anilor șaizeci. la transplanturile de piele la șoareci. Toleranța la șoarecii maturi față de grefele de piele donatoare a apărut dacă au fost injectate cu celule limfoide donatoare în perioada embrionară. Astfel de primitori, devenind maturi sexual, nu au respins grefele de piele de la donatori din aceeași linie genetică. Pentru această descoperire, Burnet și Medawar au primit Premiul Nobel în 1960.
O creștere bruscă a interesului pentru imunologie este asociată cu crearea în 1959 a teoriei selecției clonale a imunității de către F. Burnet, un cercetător care a adus o contribuție uriașă la dezvoltarea imunologiei. Conform acestei teorii, sistemul imunitar supraveghează constanța compoziției celulare a corpului și distrugerea celulelor mutante. Teoria selecției clonale a lui Burnet a stat la baza construirii de noi ipoteze și presupuneri.
În studiile lui L. A. Zilber și ale colegilor săi, efectuate în 1951-1956, a fost creată o teorie virus-imunologică a originii cancerului, conform căreia un provirus integrat în genomul celulei determină transformarea acesteia într-o celulă canceroasă.
În 1959, omul de știință englez R. Porter a studiat structura moleculară a anticorpilor și a arătat că molecula de gamma globulină constă din două lanțuri polipeptidice ușoare și două grele legate prin legături disulfurice.
Ulterior, s-a elucidat structura moleculară a anticorpilor, s-a stabilit secvența aminoacizilor din lanțurile ușoare și grele, imunoglobulinele au fost împărțite în clase și subclase și s-au obținut date importante despre proprietățile lor fizico-chimice și biologice. Pentru studiile asupra structurii moleculare a anticorpilor, R. Porter și savantul american D. Edelman au primit Premiul Nobel în 1972.
În anii 30, A. Komza a descoperit că îndepărtarea timusului duce la afectarea imunității. Totuși, adevărata semnificație a acestui organ a fost clarificată după ce savantul australian J. Miller a efectuat timectomie neonatală la șoareci în 1961, după care s-a dezvoltat un sindrom specific de deficiență imunologică, în primul rând imunitatea celulară. Numeroase studii au arătat că timusul este organul central al imunității. Interesul pentru timus a crescut în mod deosebit după descoperirea în anii 70 a hormonilor săi, precum și a limfocitelor T și B.
În 1945-1955. Au fost publicate o serie de studii în care s-a demonstrat că atunci când un organ limfoepitelial, numit punga lui Fabricius, este îndepărtat de la păsări, capacitatea de a produce anticorpi scade. Astfel, s-a dovedit că există două părți ale sistemului imunitar - dependente de timus, responsabile de reacțiile imunității celulare și dependente de punga lui Fabricius, care afectează sinteza anticorpilor. J. Miller și cercetătorul englez G. Klaman în anii 70 au arătat pentru prima dată că în reacțiile imunologice, celulele acestor două sisteme intră într-o interacțiune de cooperare între ele. Studiul cooperărilor celulare este una dintre direcțiile centrale ale imunologiei moderne.
În 1948, A. Fagreus a stabilit că anticorpii sunt sintetizați de celulele plasmatice, iar J. Gowens în 1959 a dovedit rolul limfocitelor în răspunsul imun prin transferul limfocitelor.
În 1956, Jean Dosset și colegii săi au descoperit sistemul de antigen de histocompatibilitate HLA la oameni, care a făcut posibilă efectuarea tipării țesuturilor.
Mac Devvitt a demonstrat în 1965 că genele de reactivitate imunologică (genele Ir), de care depinde capacitatea de a răspunde la antigenele străine, aparțin complexului major de histocompatibilitate. În 1974, P. Zinkernagel și R. Dougherty au arătat că antigenele complexului major de histocompatibilitate fac obiectul recunoașterii imunologice primare în reacțiile limfocitelor T la diverși antigeni.
Importantă pentru înțelegerea mecanismelor de reglare a activității celulelor imunocompetente și a interacțiunilor acestora cu celulele auxiliare a fost descoperirea în 1969 de către D. Dumond a limfokinelor produse de limfocite și crearea de către N. Jerne în 1974 a teoriei rețelei imunoregulatoare. „idiotip-antiidiotip”.
Alături de datele fundamentale obţinute, noile metode de cercetare au avut o mare importanţă pentru dezvoltarea imunologiei. Acestea includ metode pentru cultivarea limfocitelor (P. Novell), determinarea cantitativă a celulelor formatoare de anticorpi (N. Erne, A. Nordin), celulele formatoare de colonii (Mac Kulloch), metodele de cultivare a celulelor limfoide (T. Meikinodan), detectarea a receptorilor de pe membranele limfocitare. Posibilitatea utilizării metodelor de cercetare imunologică și a creșterii sensibilității acestora a crescut semnificativ datorită introducerii în practică a metodei radioimunologice. Pentru dezvoltarea acestei metode, cercetătorul american R. Yalow a fost distins cu Premiul Nobel în 1978.
Dezvoltarea imunologiei, geneticii și biologiei generale a fost foarte influențată de ipoteza exprimată în 1965 de W. Dreyer și J. Bennett că lanțul ușor al imunoglobulinelor este codificat nu de una, ci de două gene diferite. Înainte de aceasta, a fost în general acceptată ipoteza lui F Jacob și J. Monod, conform căreia sinteza fiecărei molecule proteice este codificată de o genă separată.
Următoarea etapă în dezvoltarea imunologiei a fost studiul subpopulațiilor de limfocite și hormoni timusici, care au atât efecte stimulatoare, cât și inhibitorii asupra procesului imunitar.
Dovada existenței celulelor stem în măduva osoasă capabile să se transforme în celule imunocompetente aparține perioadei ultimelor două decenii.
Realizările imunologiei din ultimii 20 de ani au confirmat ideea lui Burnet că imunitatea este un fenomen de ordin homeostatic și prin natura sa este îndreptată în primul rând împotriva celulelor mutante și auto-antigenelor care apar în organism, iar acțiunea antimicrobiană este o manifestare specială a imunitate. Astfel, imunologia infecțioasă, care se dezvoltă de mult timp ca una dintre domeniile microbiologiei, a devenit baza pentru apariția unui nou domeniu de cunoaștere științifică - imunologia neinfecțioasă.
Sarcina principală a imunologiei moderne este identificarea mecanismelor biologice ale imunogenezei la nivel celular și molecular. Se studiază structura și funcțiile celulelor limfoide, proprietățile și natura proceselor fizico-chimice care au loc pe membranele lor, în citoplasmă și organite. Ca urmare a acestor studii, astăzi imunologia s-a apropiat de înțelegerea mecanismelor intime de recunoaștere, sinteza anticorpilor, structura și funcțiile acestora. S-au făcut progrese semnificative în studiul receptorilor limfocitelor T, al cooperărilor celulare și al mecanismelor răspunsurilor imune celulare.
Dezvoltarea imunologiei a condus la identificarea unui număr de domenii independente în ea: imunologie generală, imunotoleranță, imunochimie, imunomorfologie, imunogenetică, imunologie tumorală, imunologie de transplant, imunologie de embriogeneză, procese autoimune, radioimunologie, alergii, imunobiotehnologie, imunologie de mediu, etc.
– se determină distanța de la punctul de referință la valorile specifice indicatorilor obiectelor evaluate.
În această metodă, indicatorul de evaluare integrată ia în considerare nu numai valorile absolute ale indicatorilor parțiali comparați, ci și apropierea acestora de cele mai bune valori.
Următoarea analogie matematică este propusă pentru calcularea valorii unui indicator al unei evaluări complexe a unei întreprinderi.
Fiecare întreprindere este considerată ca un punct în spațiul euclidian n-dimensional; coordonatele punctului - valorile indicatorilor prin care se realizează comparația. Este introdus conceptul de standard - o întreprindere în care toți indicatorii au cele mai bune valori dintr-un anumit set de întreprinderi. Ca standard, puteți lua și un obiect condiționat, în care toți indicatorii corespund valorilor recomandate sau standard. Cu cât întreprinderea este mai aproape de indicatorii standardului, cu atât distanța sa de punctul standard este mai mică și cu atât ratingul este mai mare. Cel mai mare rating aparține întreprinderii cu valoarea minimă a evaluării complexe.
Pentru fiecare întreprindere analizată, valoarea ratingului său este determinată de formula
unde х ij sunt coordonatele punctelor matricei - indicatorii standardizați ai întreprinderii j-a, care sunt determinați de raportul dintre valorile reale ale fiecărui indicator cu cel de referință conform formulei
X ij = a ij: a ij max
unde a ij max este valoarea de referință a indicatorului.
Este necesar să se acorde atenție validității distanțelor dintre valorile indicatorilor unui anumit obiect de studiu și standard. Aspecte separate ale activității au un impact inegal asupra situației financiare și eficienței producției. În astfel de condiții se introduc factori de ponderare; dau importanta anumitor indicatori. Pentru a obține o evaluare cuprinzătoare, ținând cont de coeficienții de greutate, utilizați formula
unde k 1 ... k n - coeficienții de pondere a indicatorilor determinați prin evaluări ale experților.
Pe baza acestei formule, valorile coordonatelor sunt pătrate și înmulțite cu coeficienții de greutate corespunzători; însumare peste coloanele matricei. Sumele subradicale rezultate sunt aranjate în ordine descrescătoare. În acest caz, scorul de rating este stabilit de distanța maximă de la originea coordonatelor, și nu de abaterea minimă de la întreprinderea de referință. Cel mai mare rating este acordat întreprinderii, care are cel mai mare rezultat total pentru toți indicatorii.
1. Rezultatele activităților financiare și economice sunt prezentate sub forma unei matrice inițiale, în care sunt evidențiate valorile de referință (cele mai bune) ale indicatorilor.
2. O matrice este compilată cu coeficienți standardizați calculați prin împărțirea fiecărui indicator real la coeficientul maxim (de referință). Valorile de referință ale indicatorilor sunt egale cu unu.
3. Se alcătuiește o nouă matrice, unde pentru fiecare întreprindere se calculează distanța de la coeficient până la punctul de referință. Valorile obținute sunt rezumate pentru fiecare întreprindere.
4. Întreprinderile sunt clasate în ordinea descrescătoare a evaluării. Cel mai mare rating este acordat întreprinderii cu cea mai mică valoare a ratingului.
PLAN
1. Definiția conceptului de „imunitate”.
2. Istoria formării imunologiei.
3. Tipuri și forme de imunitate.
4. Mecanisme de rezistență nespecifică și caracteristicile acestora.
5. Antigenii ca inductori ai antimicrobienilor dobanditi
imunitatea, natura și proprietățile lor.
6. Antigene ale microorganismelor și animalelor.
1. Definiția conceptului de „imunitate”.
Imunitate- acesta este un ansamblu de reacții și adaptări protectoare și adaptative care vizează menținerea constantă a mediului intern (homeostazia) și protejarea organismului de agenți infecțioși și alți agenți genetic străini.
Imunitatea este un fenomen biologic universal pentru toate formele organice ale materiei, multicomponent și divers în mecanismele și manifestările sale.
Cuvântul „imunitate” provine din cuvântul latin „ imunitate"- imunitate.
Din punct de vedere istoric, este strâns legat de conceptul de imunitate la agenții patogeni ai bolilor infecțioase, deoarece. doctrina imunității (imunologie) - a apărut și s-a format la sfârșitul secolului al XIX-lea în adâncurile microbiologiei, datorită cercetărilor lui Louis Pasteur, Ilya Ilyich Mechnikov, Paul Erlich și alți oameni de știință.
Introducere. Principalele etape ale dezvoltării imunologiei.
Imunologie este știința structurii și funcției sistemului imunitar al unui organism animal, inclusiv a oamenilor și a plantelor, sau știința modelelor de reactivitate imunologică a organismelor și a metodelor de utilizare a fenomenelor imunologice în diagnosticarea, terapia și prevenirea bolilor infecțioase și imune. boli.
Imunologia a apărut ca parte a microbiologiei ca urmare a aplicării practice a acesteia din urmă în tratamentul bolilor infecțioase. Prin urmare, imunologia infecțioasă s-a dezvoltat mai întâi.
De la începuturile sale, imunologia a interacționat strâns cu alte științe: genetică, fiziologie, biochimie și citologie. La sfârșitul secolului al XX-lea, a devenit o știință biologică funcțională independentă.
Există mai multe etape în dezvoltarea imunologiei:
Infecțios(L. Pasteur și alții), când a început studiul imunității la infecții. neinfectioase, după descoperirea de către K. Landsteiner a grupelor sanguine şi
fenomenul anafilaxiei de către Sh. Richet şi P. Portier.
Celular-umoral, care este asociat cu descoperirile făcute de câștigătorii premiului Nobel:
I. I. Mechnikov - a dezvoltat teoria celulară a imunității (fagocitoza), P. Ehrlich - a dezvoltat teoria umorală a imunității (1908).
F. Burnet și N. Ierne - au creat teoria modernă clonal-selectivă a imunității (1960).
P. Medawar – a descoperit natura imunologică a respingerii alogrefei (1960).
Genetica moleculara, caracterizat prin descoperiri remarcabile care au fost distinse cu Premiul Nobel:
R. Porter și D. Edelman - au decodat structura anticorpilor (1972).
C. Melstein și G. Koehler - au dezvoltat o metodă de obținere a anticorpilor monoclonali pe baza hibrizilor pe care i-au creat (1984).
S. Tonegawa - a dezvăluit mecanismele genetice ale recombinării somatice a genelor imunoglobulinei ca bază pentru formarea unei varietăți de receptori de recunoaștere a antigenelor ai limfocitelor (1987).
R. Zinkernagel și P. Dougherty - au dezvăluit rolul moleculelor MHC (complex mare de histocompatibilitate) (1996).
Jean Dosset și colegii au descoperit un sistem de antigene și leucocite umane (antigeni de histocompatibilitate) - HLA, care a făcut posibilă efectuarea tipării țesuturilor (1980).
Oamenii de știință ruși au avut o contribuție semnificativă la dezvoltarea imunologiei: I. I. Mechnikov (teoria fagocitozei), N. F. Gamaleya (vaccinuri și imunitate), A. A. Bogomolets (imunitate și alergii), V. I. Ioffe (imunitate antiinfecțioasă), P. M. Kosyakov și E. A. Zotikov (izoserologie și izoantigene), A. D. Ado și I. S. Gushchin (alergie și boli alergice),
R. V. Petrov și R. M. Khaltov (imunogenetică, interacțiune celulară, antigene artificiale și vaccinuri, noi imunomodulatori), A. A. Vorobyov (anatoxinele și imunitatea în infecții), B. F. Semenov (imunitate antiinfecțioasă), L V. Kovalchuk, B. V. Pinechin, A. N. evaluarea stării imune), N. V. Medunitsyn (vaccinuri și citotoxine), V. Ya. Arlon, A. A. Yarilin (hormoni și funcția timusului) și multe altele.
În Belarus, prima teză de doctorat în imunologie „Reacții ale imunității la transplant in vivo și in vitro în diverse sisteme imunogenetice” a fost susținută în 1974 de D. K. Novikov.
Oamenii de știință din Belarus au o anumită contribuție la dezvoltarea imunologiei: I. I. Generalov (abzime și semnificația lor clinică), N. N. Voitenyuk (citokine), E. A. Dotsenko (ecologie, astm bronșic), V. M. Kozin (imunopatologie și imunoterapie psoriazis), D. K. Novikov (imunodeficiențe). și alergii), V. I. Novikova (imunoterapia și evaluarea stării imunitare la copii), N. A. Skepyan (boli alergice), L. P. Titov (patologia sistemului complementului), M. P. Potaknev (citokine și patologie), S. V. Fedorovich (alergie profesională).
Un medic englez a fost la originile imunologiei jenner care a dezvoltat o metodă de vaccinare împotriva variolei. Cu toate acestea, cercetările sale au fost de natură privată și au vizat o singură boală.
Dezvoltarea imunologiei științifice este asociată cu numele Louis Pasteur, care a făcut primul pas către o căutare țintită a preparatelor vaccinale care să creeze imunitate stabilă la infecții: a primit și a pus în practică vaccinuri împotriva holerei, antraxului, rabiei, obținute din microbi cu virulență slăbită (atenuată).
Fondatorul teoriei imunității celulare este I.I. Mechnikov, care a creat teoria fagocitară (1901-1908).
Behring și Erlich- a pus bazele imunității umorale.
Emil von Behring- 1 Premiu Nobel pentru Medicină (1901), acordat pentru descoperirea anticorpilor anti-toxici și dezvoltarea serurilor anti-tetanos și anti-difterie.
Erlich- fondatorul teoriei lanțurilor laterale (at sub formă de receptori sunt localizați pe suprafața celulelor, ag selectează în mod specific receptorii de anticorpi corespunzători, asigură eliberarea lor în circulație și hiperproducția compensatorie de anticorpi (receptori).
Doctrina antigenelor - K. Landsteiner, J. Bordet, a demonstrat că nu numai microbii și virușii, ci și orice celule animale pot fi ag. K. Landsteiner descoperirea grupelor sanguine. (1930).
C. Richet- descoperirea anafilaxiei şi a alergiilor (1913).
Burnet și Medovar(1960) - doctrina toleranței imunologice, a arătat că aceleași mecanisme stau la baza respingerii țesuturilor străine genetic și a imunității infecțioase. M. Burnet este creatorul teoriei selecției clonale a imunității - o clonă de limfocite este capabilă să răspundă la un singur determinant antigenic specific. Și în plus, Burnet este autorul uneia dintre cele mai importante prevederi ale imunologiei - conceptul de supraveghere imunologică a constanței mediului intern al organismului.
În anii 60, doctrina sistemelor de imunitate T și B a început să se dezvolte rapid ( Claman, Davis, Roit).
A fost propusă o teorie a cooperării cu 3 celule a imunocitelor în răspunsul imun ( Petrov, Royt si etc.). Principalii participanți la schema propusă au fost limfocitele T și B și macrofagele.
descifrarea structurii Ig - ( Porter, Adelman)
descoperirea structurilor codificate de MHC - ( Benaceraf, Snell)
controlul genic al răspunsului imun, diversitatea anticorpilor și importanța anumitor gene în susceptibilitatea la boli
Obținerea anticorpilor monoclonali și fundamentarea reglării rețelei a imunogenezei ( Koehler, Milstein, Jerne)
În prezent, există o dezvoltare intensivă a imunologiei clinice și introducerea pe scară largă în medicina practică a realizărilor imunologiei teoretice (descifrarea patogenezei multor boli; crearea de noi clasificări; clasificarea bolilor sistemului imunitar; dezvoltarea metodelor de imunodiagnostic (ELISA, RIA, reacție în lanț a polimerazei etc.), imunoterapie) .
Principalele etape ale formării și dezvoltării imunologiei:
1796 - 1900– imunologie infecțioasă
1900 - 1950- imunologie normală
1950 până în prezent- scena modernă
Scurtă schiță istorică a dezvoltării imunologiei
Lumea antică și Evul Mediu
1000 î.Hr - primele inoculări ale conținutului de papule de variolă la oameni sănătoși pentru a le proteja de forma acută a bolii au fost efectuate în China, apoi s-au răspândit în India, Europa, Asia Mică și Caucaz.
Primele vaccinuri
Din 1701, variolația (vaccinarea împotriva variolei) s-a răspândit la Constantinopol, de unde se răspândește în Europa. În 1722, prințul și prințesa de Wales le-au inoculat două dintre fiicele lor cu variolă, oferind un exemplu regal pentru poporul Angliei. La Londra, în 1746, a fost deschis un spital special din St. Pancras, în care variola era inoculată tuturor. La 12 octombrie 1768, unul dintre cei mai buni inoculatori, Thomas Dimsdale, i-a vaccinat pe împărăteasa Ecaterina a II-a și pe fiul ei Paul. În 1796, după treizeci de ani de cercetări, Edward Jenner a testat metoda de inoculare a oamenilor cu variola bovină pe un băiețel de 8 ani, iar apoi pe încă 23 de persoane. În 1798 a publicat rezultatele cercetărilor sale. Jenner a dezvoltat o tehnică medicală pentru vaccinarea împotriva variolei, pe care a numit-o vaccinare (din latină vaccus - vaca).
Revoluție imunologică
În 1880, Louis Pasteur a publicat un articol despre protejarea găinilor de holeră prin imunizarea lor cu un agent patogen cu virulență redusă.
În 1881, Pasteur a efectuat un experiment public în care 27 de oi au fost vaccinate cu un vaccin cu antrax, iar în 1885 a testat cu succes vaccinul antirabic pe un băiat mușcat de un câine turbat. Aceste evenimente marchează nașterea imunologiei infecțioase și începutul erei vaccinării. În 1890, medicul german Emil von Behring, împreună cu Shibasaburo Kitasato, a arătat că în sângele persoanelor care au avut difterie sau tetanos se formează antitoxine, care asigură imunitate la aceste boli atât celor care au fost bolnavi, cât și celor cărora le-au fost. astfel de sânge va fi transfuzat. În același an, pe baza acestor descoperiri, a fost dezvoltată o metodă de tratament cu ser sanguin. Lucrările acestor oameni de știință au marcat începutul studiului mecanismelor imunității umorale. În 1883, biologul și imunologul rus Ilya Mechnikov a făcut primul raport despre teoria fagocitară a imunității. Mechnikov a fost cel care a stat la originile cunoașterii problemelor imunității celulare. Mechnikov a arătat că în corpul uman există celule mobile speciale ameboide - neutrofile și macrofage care absorb și digeră microorganismele patogene. Lor le-a dat rolul principal în protejarea corpului.
În 1891, a fost publicat un articol de Paul Ehrlich, în care a folosit termenul „anticorp” pentru a se referi la substanțele antimicrobiene din sânge. În paralel cu Mechnikov, Erlich și-a dezvoltat teoria apărării imune a organismului. Ehrlich a remarcat că principala proprietate a anticorpilor este specificitatea lor pronunțată. Două teorii - fagocitară (celulară) și umorală - în perioada apariției lor au stat pe poziții antagonice. În 1908, Mechnikov și Erlich au împărțit Premiul Nobel pentru Medicină, iar mai târziu s-a dovedit că teoriile lor se completează reciproc.
În 1900, imunologul austriac Karl Landsteiner a descoperit grupele sanguine umane.În 1904, celebrul chimist Svante Arrhenius a dovedit reversibilitatea interacțiunii antigen-anticorp și a pus bazele imunochimiei. În 1913, a fost organizată Asociația Americană a Imunologilor. Revoluție în imunologia teoretică Virologul Frank Macfarlane Burnet a devenit autorul teoriei clonale selective a imunității și descoperitorul fenomenului de imunotoleranță.
Studiul imunoglobulinelor a început cu lucrările din 1937 asupra electroforezei proteinelor din sânge de Arne Tiselius. Apoi în anii 40-60. au fost descoperite clase și izotipuri de imunoglobuline, iar în 1962 Rodney Porter a propus un model al structurii moleculelor de imunoglobuline, care s-a dovedit a fi universal pentru imunoglobulinele de toate izotipurile și este absolut corect până în ziua de azi în cunoștințele noastre.
Anii 60 - începutul anilor 80 - etapa de izolare a diverșilor factori - mediatori umorali ai răspunsului imun din supernatanții culturii celulare. De la mijlocul anilor 1980 până în prezent, metodele de clonare moleculară, șoarecii transgenici și șoarecii cu îndepărtarea genelor specificate (knokout) au intrat în imunologie.
În lucrările lui James Govans din anii 60 ai secolului XX. se arată rolul limfocitelor în organism. La mijlocul secolului XX. o echipă condusă de geneticianul și imunologul american George Snell a efectuat experimente cu șoareci care au dus la descoperirea complexului major de histocompatibilitate și a legilor transplantului.
În 2011, imunologul francez Jules Hoffmann a primit Premiul Nobel pentru Fiziologie sau Medicină pentru munca sa „pentru studiul activării imunității înnăscute”.
În secolul XXI, principalele sarcini ale imunologiei au devenit: studiul mecanismelor moleculare ale imunității, atât înnăscute, cât și dobândite, dezvoltarea de noi vaccinuri și metode de tratare a alergiilor, imunodeficiențelor și a bolilor oncologice.
Subiectul, scopurile și obiectivele imunologiei
În funcție de metoda și obiectul cunoașterii, imunologia poate fi împărțită în generală și particulară. Imunologia generală studiază procesele de „imunitate la nivel molecular, celular și organismal, genetica și evoluția imunității, reglarea imunității la toate nivelurile. Imunologia privată studiază metode și metode pentru prevenirea, diagnosticarea și tratamentul bolilor infecțioase (imunoprofilaxie, vaccinologie). ); tumori maligne (imuno-oncologie); afecțiuni propice transplantului de organe și țesuturi străine (imunologie de transplant), reacții perverse la antigene (alergologie, imunopatologie), influență a factorilor de mediu asupra sistemului imunitar (imunologie de mediu).
Sarcini de imunologie:
1. studiul sistemului imunitar al unei persoane sănătoase;
2. studiul rolului IP în patogeneza bolilor infecțioase și netransmisibile
3. dezvoltarea unor metode unificate şi informative de evaluare a stării imune
4. dezvoltarea de noi medicamente imunoactive de mare eficiență și scheme optime de utilizare a acestora.
subiectul principal cercetarea în imunologie este cunoașterea mecanismelor de formare a unui răspuns imun specific al organismului la toți compușii străini și antigenici.
Cele mai caracteristice trăsături ale sistemului imunitar, care îl deosebesc de alte sisteme ale corpului, sunt următoarele:
1. Capacitatea de a diferenția totul „propriu” de tot ceea ce „străin”;
2. Crearea memoriei din contactul primar cu material antigenic străin;
3. Organizarea clonală a celulelor imunocompetente, care se manifestă prin capacitatea unei singure clone de celule de a răspunde doar la unul dintre numeroșii determinanți antigenici.
Caracteristicile generale ale sistemului imunitar al mamiferelor
Organele sistemului imunitar sunt de obicei împărțite în centrale (sau primare) și periferice (sau secundare), în funcție nu atât de localizarea lor în organism, cât de gradul de importanță al acestora în menținerea stării normale a acestui sistem. Măduva osoasă roșie și timusul (glanda timus) sunt clasificate drept organe primare ale sistemului imunitar datorită faptului că în ele apar și trec prin principalele etape de dezvoltare celulele care alcătuiesc sistemul imunitar. Sunt considerate secundare acele organe în care aceste celule desfășoară doar unele etape ale dezvoltării lor și sunt localizate temporar în cursul circulației inerente acestor celule în întregul corp. Acestea din sistemul imunitar sunt splina, ganglionii limfatici și acumulările limfoide neseparate de țesuturile înconjurătoare prin membranele țesutului conjunctiv: amigdalele și adenoidele nazofaringelui, precum și formațiunile limfoide specifice din pereții intestinali, numite plasturi Peyer.
Sistemul imunitar, datorită mobilității celulelor sale constitutive, este distribuit în tot organismul. Celulele la care se face referire, inițial celule sanguine, sunt capabile să pătrundă în pereții capilarelor și să se deplaseze între celulele altor țesuturi, ceea ce face ca mediul intern practic oriunde în organism să fie accesibil sistemului imunitar. Mai exact, celulele sistemului imunitar sunt considerate a fi toate leucocite din sânge, împărțite condiționat în 5 grupe: monocite, neutrofile, eozinofile, bazofile și limfocite. În condiții fiziologice normale, bazofilele (după pătrunderea în țesut se numesc mastocite) și monocitele, care se transformă în așa-numitele macrofage tisulare în timpul unor astfel de mișcări, au capacitatea de a se muta din sânge în țesuturi. În organele limfoide secundare, limfocitele sunt, de asemenea, capabile să treacă din sânge în țesuturi, dintre care unele se pot întoarce din nou în fluxul sanguin. Limfocitele sunt de obicei împărțite în funcție de locurile formării lor primare în limfocite T (trec prin principalele stadii de maturare în timus) și limfocite B (la mamifere se maturizează în principal în măduva osoasă roșie).
A treia componentă a sistemului imunitar sunt moleculele secretate de celulele sale, deoarece unele dintre ele sunt capabile să funcționeze ca agenți cu acțiune proprie în timpul implementării reacțiilor de protecție. Un exemplu tipic de astfel de molecule sunt imunoglobulinele secretate de limfocitele B (numite și anticorpi), care pot interacționa în mod specific cu antigeni străini specifici fără nicio influență a altor componente ale sistemului imunitar. Pe lângă imunoglobuline, moleculele inerente sistemului imunitar sunt considerate a fi substanțe care reglează activitatea atât a celulelor sistemului imunitar, cât și a altor celule ale organismului, cel mai adesea sunt numite: citokine, limfokine și interleukine.
Structura și caracteristicile organelor centrale și periferice ale sistemului imunitar
măduvă osoasă (centrală) localizat în cavitatea internă a oaselor tubulare și este o unire tisulară a stromei reticulare, celule hematopoietice și limfoide dens împachetate, precum și o rețea extinsă de capilare.Scopul principal este producerea de celule sanguine și limfocite. Dezvoltarea elementelor celulare ale măduvei osoase începe de la celula stem hematopoietică (HSC), care dă naștere la șase muguri de diferențiere:
1) megacariocitar, care se termină cu formarea trombocitelor;
2) eritroid, cu formarea de globule roșii nenucleare, purtătoare de oxigen; 3) granulocitare, din care se formează: bazofile, eozinofile, neutrofile; aceste celule sunt direct implicate în procesele de inflamație și fagocitoză și participă la o formă de protecție împotriva agenților patogeni; 4) monocite-macrofag-formarea de monocite care migrează în sânge; forme finale mature - macrofagele tisulare sunt localizate în diverse organe și țesuturi;
5) formarea celulelor T a precursorului celulelor T;
6) celula B; Diferențierea celulelor B este caracterizată de finalizarea aproape completă.
timus(glanda timus) - un organ limfoepitelial situat la majoritatea mamiferelor în partea superioară a cavității toracice deasupra inimii; este format din doi lobi, împărțiți în lobuli mai mici. Organul în ansamblu și lobulii individuali sunt închise într-o capsulă de țesut conjunctiv, a cărei cavitate internă include o rețea epitelială umplută cu limfocite (timocite). Lobul - două straturi: cortexul cu o împachetare densă de timocite mici și medulara (stratul medular), unde numărul de timocite este redus.
Particularitatea organizării timusului este prezența a două unități structurale-histologice elementare: foliculii lui Clark
(parcă „cărămizi” separate din care este construit stratul cortical; limfocitele și macrofagele dens împachetate și celulele dendritice situate printre ele sunt înconjurate de celule epiteliale, care împreună creează o unitate structural-histologică elementară) și Corpul lui Hassall
(în zona medulară, acumulări rotunjite de celule epiteliale libere de limfocite; scopul funcțional al corpurilor este neclar).
Pungă de Fabricius în păsăriîndeplinește rolul de organ central al imunității, fiind furnizor de celule B pentru periferie, este locul de formare activă a producătorilor de anticorpi. Acesta este un organ limfoepitelial situat în partea din spate a cloacii. Lumenul bursei este căptușit cu epiteliu columnar. În spatele stratului epitelial se află noduli (lobuli). Cortexul este reprezentat în principal de o acumulare densă de limfocite mici. Medula mai ușoară include limfocite mari, celule plasmatice, macrofage, granulocite și celule reticulare.
Splina (periferică)- un organ mare situat în partea superioară, stângă a peritoneului. Din exterior, organul este înconjurat de o capsulă de țesut conjunctiv, din care se extind în organ pereții de susținere, trabecule. O trăsătură caracteristică a structurii este prezența a două zone - roșie (localizarea unui număr mare de eritrocite, precum și macrofage, megacariocite, granulocite, limfocite) și pulpă albă (acumulare de limfocite în jurul unui canal arterial situat excentric). Nu există granițe clare între pulpa albă și roșie, iar între ele are loc un schimb celular parțial. Limfocitele T și B sunt localizate în pulpa albă. Celulele T sunt situate în jurul arteriolelor, formând ambreiaje periarteriale. Celulele B fac parte din centrele germinale, care sunt situate în zona de graniță. Pulpa roșie conține, de asemenea, limfocite și celule plasmatice. Cu toate acestea, ei nu formează grupuri formate morfologic în această zonă.
Ganglionii limfatici sunt adevărate formațiuni limfoide. Sunt situate sub formă de boabe de-a lungul vaselor limfatice; se formează ca urmare a acumulării de celule mezenchimale în jurul vaselor de sânge. Stratul exterior al mezenchimului se diferențiază într-o capsulă de țesut conjunctiv, din care partițiile se extind în nod. Direct sub capsulă se află sinusul marginal, unde limfa pătrunde prin vasele care aduc limfa. Din sinusul marginal, limfa pătrunde în sinusurile intermediare, pătrunzând pe toată grosimea ganglionului, și este colectată în vasul limfatic care o duce în ductul toracic. Punctul de ieșire al vasului se numește poarta nodului. Vasele de sânge trec prin poartă în nod. În ganglion limfatic se disting un strat cortical și o medulară situată în centrul ganglionului. Stratul cortical al nodului este locul de concentrare a celulelor B. Medula este reprezentată de limfocite relativ slab împachetate, celule plasmatice, macrofage libere și celule stromale reticulare. Zona dintre cortex și medular este locul de concentrare a celulelor T.
Țesut limfoid localizat în pereții tractului digestiv, respirator și urogenital. Este denumit țesut limfoid asociat cu membranele mucoase. Țesutul se prezintă fie sub formă de infiltrație difuză, fie sub formă de acumulări nodulare, lipsite de carcasă de țesut conjunctiv închis. Functii: concentrează antigenul, asigură contactul cu antigenul diferitelor tipuri de celule, transportă structurile celulare ale țesutului limfoid în părțile necesare ale corpului și elimină antigenele străine. Există țesut limfoid lax – în care domină fibrele reticulare, celulele reticulare și macrofagele fixe; și dens - limfocite, plasmocite și macrofage libere.
Conceptul de imunitate. imunitatea naturală. Forme active și pasive de imunitate.
Imunitatea este imunitatea organismului la bolile infecțioase, precum și la agenți și substanțe care au proprietăți antigenice străine organismului.
Reacțiile imune sunt de natură protectoare, adaptativă și au ca scop eliberarea organismului de antigenele străine care intră în el din exterior și încalcă constanța mediului său intern. De natură defensivă, reacțiile imune, dintr-un motiv sau altul, pot fi pervertite și direcționate către unele componente proprii, normale, nemodificate ale celulelor și țesuturilor, ducând la boli autoimune. Reacțiile imune pot determina creșterea sensibilității organismului la anumiți antigeni - alergii, anafilaxie. Există următoarele tipuri de imunitate : Natural și artificial. imunitatea naturală pot fi congenitale sau dobândite. Cu imunitate naturală înnăscută, o persoană este imună de la naștere la o anumită boală. Dobândit natural numită imunitate, care apare după transferul oricărei boli infecțioase. Copiii care au avut rujeolă, oreion, tuse convulsivă dobândesc imunitate naturală împotriva acestor boli, adică nu se îmbolnăvesc din nou. În sângele unei persoane, după infectarea cu agenți patogeni ai unei boli, apar substanțe speciale de protecție, care se numesc anticorpi sau substanțe imunitare. Ei fie distrug agenții cauzali ai acestei boli, fie le slăbesc brusc acțiunea, ceea ce creează condiții favorabile pentru fagocitoză. Imunitatea naturală dobândită durează luni sau ani.
Activ imunitatea naturală dobândită apare după o boală infecțioasă. Aceasta este cea mai durabilă, imunitate de lungă durată, care se menține uneori pe tot parcursul vieții. Imunitatea artificială dobândită activ rezultă din vaccinarea cu vaccinuri vii atenuate sau ucise (microbiene). O astfel de imunitate apare la 1-2 săptămâni după vaccinare și se menține o perioadă relativ lungă de timp - ani și zeci de ani. Pasiv imunitatea naturală dobândită este imunitatea fătului sau a nou-născutului care primește anticorpi de la mamă prin placentă sau laptele matern. Imunitatea artificială dobândită pasiv este creată prin introducerea în organism a imunoglobulinelor obținute de la persoane sau animale imunizate activ. O astfel de imunitate se stabilește rapid - la câteva ore după introducerea serului imun sau a imunoglobulinei și persistă pentru o perioadă scurtă de timp timp de 3-4 săptămâni, deoarece organismul caută să scape de serul străin. Toate tipurile de imunitate asociate cu formarea de anticorpi sunt numite specific, deoarece anticorpii acționează numai împotriva unui anumit tip de microorganisme sau toxine.
La nespecifice mecanismele de protecție includ pielea și membranele mucoase, care sunt practic impermeabile la microbi, lizozima (o substanță bactericidă a pielii și a membranelor mucoase), o reacție inflamatorie, proprietățile bactericide ale sângelui fluidului tisular și reacții de fagocitoză.
Imunitatea artificială și rolul ei în lupta împotriva bolilor infecțioase. Conceptul de vaccinuri și seruri utilizate pentru prevenirea bolilor infecțioase
Imunitatea artificială este imunitatea care este creată ca urmare a activării sistemului imunitar sau a imunizării artificiale. Există imunitate artificială pasivă și activă. Imunitatea pasivă apare datorită introducerii în organism a unor seruri specifice, interferoni și amestecurile acestora, interleukine, imunoglobuline, celule de măduvă osoasă, monocite, limfocite, care sunt activate artificial in vitro. Imunitatea pasivă este creată cu imunodeficiența secundară primară sau severă. Imunitatea activă este creată prin activarea mecanismelor de răspuns imun. Pentru aceasta se folosesc vaccinuri, inductori de interleukine, interferoni, activatori ai fagocitozei și sisteme de complement, mecanisme natural killer. Cu imunizarea activă, organismul însuși produce interferoni, anticorpi, interleukine și alți factori de imunitate. Vaccinul conține viruși sau bacterii slăbiți sau uciși. Se dezvoltă un răspuns imun primar, iar după ce intră un agent patogen neslăbit, este oferit și un răspuns secundar, care contribuie la o evoluție ușoară a bolii și la o recuperare rapidă.
Vaccinurile și serurile sunt utilizate ca imunostimulatori activi sau pasivi. Astfel de medicamente sunt deosebit de eficiente dacă sunt utilizate nu numai pentru tratament, ci și pentru prevenirea bolilor infecțioase. Vaccinuri
sunt produse direct din microorganismele care provoacă infecții sau din antigenele acestora. Vaccinul ajută organismul să producă singur anticorpi pentru a lupta împotriva virușilor sau infecțiilor.În funcție de originea vaccinului, aceștia se împart în:
vaccinuri corpusculare (astfel de medicamente sunt produse din microbii uciși care provoacă boala),
vaccinuri atenuate (produse din microorganisme slăbite),
vaccinuri chimice în care antigenele sunt create chimic într-un laborator (în special, vaccinurile împotriva hepatitei B).
Seruri sunt plasmă sanguină fără fibrinogen. Serul se obține prin coagularea naturală a plasmei sau cu ajutorul ionilor de calciu, care precipită fibrinogenul. Odată cu introducerea serului are loc și formarea sistemului imunitar. Serul este de obicei făcut din sânge animal, dar cel mai eficient în unele cazuri este serul pe bază de sânge uman - imunoglobuline (sau gama globuline). γ-globulinele nu provoacă reacții alergice. Serurile conțin anticorpi gata preparate, care sunt utilizați dacă organismul nu îi poate produce singur din cauza imunodeficienței severe, pentru tratamentul și prevenirea infecțiilor virale sau bacteriene (dar nu în formă acută). Serurile pot fi folosite după transplantul de organe pentru a preveni posibila respingere a acestora de către organism. Serurile sunt, de asemenea, folosite pentru a forma imunitatea unei persoane la infecții dacă trebuie să intre în contact cu persoane care sunt deja bolnave sau purtători ai anumitor viruși.
Mecanisme de apărare constitutive și inductibile ale organismului mamifer împotriva infecției.
Trăsături distinctive ale constitutive (congenitale)
scut mecanismele sunt prezența lor constantă în organism
indiferent de acţiunea factorilor destabilizatori şi de absenţa
specificitate pronunțată, adică asemănarea manifestării sub acțiune
diverși factori. Acest tip de mecanisme de apărare sunt capabile
proteja temporar organismul de o serie de factori aproape imediat
la scurt timp după naștere. În același timp reacții de apărare inductibile
absente în organism inițial, apar în timpul vieții în re-
ca urmare a contactului cu un anumit factor destabilizator şi zona
dați o specificitate pronunțată, adică protejați numai de
factor care a determinat manifestarea acestui mecanism.
Se poate considera că mecanisme constitutive de apărare sunt prima barieră sau eșalon de apărare împotriva agresiunii biologice și inductibil - al doilea, deoarece acestea, de regulă, pornesc numai atunci când prima barieră este depășită într-o măsură sau alta.
La constitutiv bariere de protecție în mod tradițional referi nu-
permeabilitatea tegumentului, lizozima, enzimele hidrolitice și
acid clorhidric al tractului gastrointestinal, interferon, inflamator
ion, fagocitoză, sistemul complementului și altele prezente în
factori umorali sanguini de protectie constitutiva.
Mecanismele de apărare inductibile sunt toate formele de imunitate
răspuns bazat pe recunoașterea specifică a anti-
genele. De regulă, implementarea lor necesită mult mai mult timp.
nimic pentru manifestarea factorilor constitutivi de protecție, precum și obligatorii
participarea celulelor imunocompetente este esențială. De bază și
Cele mai studiate dintre ele sunt: răspuns la dependenta de timus
antigene, ducand la aparitia anticorpilor specifici si corespunzatori
ramificarea celulelor memoriei imune; acţiunea T-killers, limitând
redus de molecule ale complexului major de histocompatibilitate;
hipersensibilitate de tip întârziat; hipersensibilitate
tip imediat.
Funcția de protecție a pielii și mucoaselor mamiferelor.
Pielea însăși (derma) reprezentată de un fibros dens
țesut unificator, al cărui semn distinctiv este Disponibilitate
o cantitate mare de substanță intercelulară densă. Principal
componentele acestei substanțe sunt proteinele de colagen și elastina, care se formează
fibre vâscoase și umplerea spațiului dintre aceste fibre
acid hialuronic polizaharid. Această combinație creează un puternic
strâns și în același timp barieră mecanică de întindere pe un drum
cautand sa patrunda in microorganisme. Disponibil pe piele glandele sudoripare pe lângă îndeplinirea principalului său functie termostatica joacă un rol important în formarea proprietăților protectoare ale pielii. Prezența unor cantități mici de compuși organici cu greutate moleculară mică (acid lactic, unii aminoacizi, acid uric și uree) în lichidul transpirator și aciditatea acestuia (pH 5,5) sunt un factor nefavorabil pentru bacterii și ciuperci. Acțiunea combinată a acestor secrete în general face suprafața pielii bactericidă sv-va, ceea ce este confirmat experimental prin moartea bacteriilor saprotrofe plasate pe suprafața pielii curate în decurs de 1 oră de la aplicare. De asemenea, ar trebui să sublinieze importanța secreției glandelor sebacee ca un hidrofug deoarece microorganismele care ajung la suprafața pielii cu apă (de exemplu, atunci când înotați în rezervoare naturale) sunt îndepărtate atunci când apa se scurge din pielea neumedată. În același timp, aceeași secreție grasă protejează pielea de uscăciune și ulterior cracare, ceea ce i-ar reduce drastic protecția. Membranele mucoase asigură protecţie corpul într-un mod diferit. Datorită absenței aproape complete a spațiilor intercelulare în compoziția țesuturilor epiteliale care formează mucoasele. ve-va rezistenţa mecanică a mucoaselor este extrem de scăzută iar celulele mucoasei sunt destul de usor deteriorate de factori externi. Cu toate acestea, capacitatea lor mare de regenerare face posibilă compensarea
repara daunele apărute și stratul secretat de aceste celule
slime previne direct efectul micro-mișcărilor asupra celulelor. Îndepărtarea definitivă a secretelor alocate ca urmare a trecerii
drenaj puternic sau activitate prezentă în unele mucoase
cochilii de celule ciliare contribuie şi îndepărtarea celor prinși
suprafața particulelor. Deoarece procesul de astfel de îndepărtare, de regulă, este prelungit în timp, majoritatea secrețiilor mucoase conțin substanțe bactericide. Acest lucru este cel mai pronunțat în membranele mucoase. tractului respirator și ochilor, unde este prezentă compoziția mucusului secretat.înseamnă. număr lizozim-acetilmuramidaza, substrat
pentru care yavl.una din componentele principale ale custii. ziduri
bacterii - peptidoglican mureina. În plus, prezent în mucus
nas polizaharidele au unele antivirusuri
acțiune.
Rolul microflorei umane normale în protecția împotriva infecțiilor.
Microflora normală joacă un rol important în protejarea organismului de microbii patogeni, de exemplu prin stimularea sistemului imunitar, participând la reacțiile metabolice. În același timp, această floră poate duce la dezvoltarea bolilor infecțioase. Rolul microflorei normale în infecții Majoritatea infecțiilor cauzată de reprezentanții microflorei normale este de natură oportunistă. În special, anaerobii intestinali (de exemplu, bacteroides) pot provoca formarea de abces după pătrunderea în peretele intestinal ca urmare a unui traumatism sau a unei intervenții chirurgicale; Principalii agenți cauzali ai pneumoniei post-gripale frecvent înregistrate sunt micro-noi, care trăiesc în nazofaringele oricărei persoane. Numărul unor astfel de leziuni este atât de mare încât se pare că medicii sunt mai predispuși să se ocupe de infecții endogene decât exogene, adică de patologia indusă de microflora endogenă. Lipsa unei distincții clare între microbii oportunişti iar comensalii sugerează că colonizarea nerestricționată de către orice tip de bacterie care poate supraviețui în corpul uman poate duce la dezvoltarea unei patologii infecțioase. Dar această poziție este relativă - diferiți membri ai comunităților microbiene prezintă proprietăți patogene de diferite ordine (unele bacterii sunt mai susceptibile de a provoca leziuni decât altele). De exemplu, în ciuda diversității microflorei intestinale, peritonita cauzată de o pătrundere a bacteriilor în cavitatea abdominală este cauzată doar de câteva tipuri de bacterii. Rolul principal în dezvoltarea unor astfel de leziuni este jucat nu de virulența agentului patogen în sine, ci de starea sistemelor de protecție ale macroorganismului; Astfel, la persoanele cu imunodeficiență, microorganismele slab virulente sau avirulente (candida, pneumocystis) pot provoca leziuni severe, adesea fatale. Microflora normală este concurenta pentru patogen; mecanismele de inhibare a creșterii acestora din urmă sunt destul de diverse. Mecanismul principal- legarea selectivă de către microflora normală a receptorilor de suprafață ai celulelor, în special a celor epiteliali. Aceste proprietăți sunt deosebit de pronunțate la bifidobacterii și lactobacili; potentialul antibacterian se formeaza prin secretia de acizi, alcooli, lizozime, bacteriocine si alte substante. Microflora normala - stimulent nespecific("iritant") al sistemului imunitar; absența biocenozei microbiene normale provoacă numeroase tulburări ale sistemului imunitar. Microflora intestinală normală joacă un rol importantîn metabolic procesele corpului și menținerea echilibrului acestora. Bacteriile intestinale sunt implicate în inactivare produse toxice de origine endo- și exogenă. Acizii și gazele eliberate în timpul vieții microbilor intestinali au un efect benefic asupra motilității intestinale și golirea acestuia în timp util.
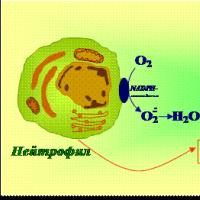
Dezvoltarea și caracterizarea celulelor fagocitare de mamifere
fagocite- celule ale sistemului imunitar care protejează organismul prin înghițirea (fagocitoză) particulelor străine dăunătoare, bacteriilor și celulelor moarte sau pe moarte. Principalele celule fagocitare ale organismului mamiferelor sunt împărțite în micro- și macrofage.
Monoblastele, sub influența unor factori umorali precum factorul de stimulare a coloniilor monocite-macrofage (M-CSF) și parțial interleukina-6 (IL-6), se transformă în promonocite, iar cele în monocite. Această etapă de dezvoltare are o durată medie de 50-60 de ore, dar monocitele intră în sânge după alte 13-26 de ore. Se crede că monocitele sunt direct în sânge timp de cel mult 4 zile, iar cele mai multe dintre ele deja în a doua zi se deplasează prin pereții capilarelor, transformându-se în macrofage tisulare. Durata de viață a macrofagelor variază în funcție de locul în care sunt localizate, dar în majoritatea cazurilor ele există timp de aproximativ 40 de zile. Macrofagele mature se disting prin prezența pe suprafața lor a unor molecule specifice necesare manifestării funcțiilor caracteristice macrofagelor. Deoarece una dintre funcțiile lor principale este fagocitoza, macrofagele au receptori care leagă lipopolizaharidele bacteriene, dintre care cea mai pronunțată este molecula CD14. O trăsătură distinctivă a macrofagelor este capacitatea lor de a se mișca activ, care se datorează proprietăților speciale ale citoscheletului lor și prezenței pe suprafața lor a unui alt grup de molecule specializate - receptorii de chemokine. Principalele celule fagocitare dintre microfage sunt neutrofile- cel mai numeros grup dintre toate leucocite, la un adult sănătos, numărul acestora este de aproximativ 70% din numărul total de globule albe. Speranța lor de viață nu este lungă - 2-3 zile, iar după ce părăsesc măduva osoasă roșie, ei rămân în sânge doar 8-10 ore, apoi se mută în țesuturi, unde mor fie în procesul de luptă împotriva agenților străini, fie prin apoptoză. Eozinofileîn organism este mult mai puțin - de la 0,5 la 2% din numărul total de leucocite. Ele se dezvoltă în mod similar cu neutrofilele, dar dezvoltarea lor este cea mai sensibilă la IL-5, cunoscută ca factor de creștere și diferențiere a eozinofilelor. Bazofilele sunt cel mai mic grup de granulocite - numărul lor la mamifere este estimat la 0,2-0,5% din numărul total de leucocite. Acestea sunt celule foarte granulare cu granule colorate cu coloranți bazici cu conținut diferit. Transformarea bazofilelor în mastocite are loc datorită pătrunderii primelor prin pereții capilarelor atât în organele limfoide secundare, cât și în epiteliul aflat în contact cu mediul și straturile subiacente ale acestuia, sau în pielea însăși. Mastocitele sunt mari în comparație cu bazofilele, numărul de granule crește în ele, iar suprafața lor capătă o structură viloasă.
Procesul de fagocitoză. Mecanisme de inactivare a microorganismelor de către fagocite. Fagocitoza incompletă, semnificația sa în dezvoltarea procesului infecțios
În mod convențional, întregul proces este de obicei împărțit în mai multe etape. Prima dintre acestea este mișcarea chemotactică a celulei fagocitare către obiectul fagocitozei. Atractanții pentru fagocite pot fi atât substanțe secretate de un agent străin care a pătruns în mediul intern, cât și substanțe care au apărut în fluidul tisular ca urmare a impactului unui agent străin asupra celulelor corpului. În special, atunci când celulele bacteriene sunt distruse, în fluidul tisular apare o peptidă scurtă constând din formil-metionină, leucină și fenilalanină, care este inițiatorul sintezei proteinelor la procariote și este absolut necaracteristic celulelor eucariote. Printre cei mai tipici chemoatractanți de origine proprie se numără mediatorii inflamatori, produșii de activare ai sistemului complementului (C3a și C5a), substanțele formate în timpul începerii sistemului de coagulare a sângelui (trombina, fibrina) și citokinele secretate de diferite celule sanguine. Pentru aceste substanțe, pe suprafața celulelor fagocitare există receptori specifici, adăugarea unui agent activ la care provoacă o modificare a proteinei G asociate receptorilor, ceea ce duce la declanșarea unui număr de procese. În special, susceptibilitatea celulelor la diferite tipuri de factori de activare crește, activitatea secretorie a fagocitelor crește, dar principalul lucru în legătură cu chemotaxia este rearanjarea citoscheletului și, în consecință, polarizarea celulelor. Celula se întoarce de la rotundă la triunghiulară, în partea citoplasmei orientată spre direcția de mișcare, numărul de organele scade și apare o rețea de microfilamente formată din F-actină, a cărei contracție determină mișcarea întregii celule în Direcția corectă. Pe membrana din această parte a celulei, integrinele apar în număr mai mare - molecule specifice pentru a îmbunătăți aderența unei celule în mișcare la pereții capilarelor sistemului circulator și producerea de catepsine, colagenază și elastază de către fagocit, care favorizează pătrunderea prin membranele bazale de la baza epiteliului, este, de asemenea, îmbunătățită. Tocmai datorită unor astfel de modificări, celulele fagocitare se pot deplasa rapid de la sânge la locul leziunii tisulare, adică potențiala penetrare a agenților străini. Unele microorganisme patogene au dobândit, în cursul evoluției comune cu gazda, capacitatea de a rezista efectelor inactivatoare ale fagocitelor și de a menține viabilitatea în timp ce se află în fagolizozomi - fagocitoză incompletă. Mecanismele care contribuie la această supraviețuire variază în funcție de speciile patogene, dar s-a demonstrat în mod clar că unele bacterii sunt capabile să producă catalază, reducând astfel efectul bactericid al căilor de inactivare dependente de oxigen.
Caracterizarea inflamației ca reacție de protecție a organismului
Inflamația este o reacție locală de protecție și de adaptare a întregului organism care apare ca răspuns la impactul unui agent dăunător. Inflamația este protejată de efectele factorilor nocivi sub forma formării unui fel de barieră. Datorită reacției inflamatorii, focarul de afectare este delimitat de întregul organism; celulele albe din sânge se grăbesc la el, efectuând fagocitoză. Inflamația include trei componente cele mai importante: alterare - o schimbare până la deteriorarea celulelor și țesuturilor, exudația - eliberarea de lichid și celule sanguine din vase și proliferarea - reproducerea celulelor și creșterea țesuturilor. În funcție de predominanța uneia dintre ele, există trei forme principale de inflamație: alternativă, exsudativă și proliferativă. Alternativă - când predomină afectarea celulelor, aceasta apare mai des în inimă, ficat, rinichi. Inflamația exudativă - odată cu aceasta, predomină modificările vaselor în focarul inflamației, ceea ce duce la o creștere bruscă a permeabilității pereților vaselor, partea lichidă a sângelui și leucocitele părăsesc vasele în țesutul din jur; lichidul care se acumulează în focar se numește exudat. Proliferativ - caracterizat prin predominanța reproducerii elementelor celulare, care se manifestă prin formarea de noduli (granuloame), îngroșări în țesut.
Sistemul de complement, modalitățile de activare și mecanismul de acțiune al acestuia.
Complementul este un termen colectiv pentru un sistem de aproximativ 20 de proteine, dintre care multe sunt precursori de enzime (proenzime). Principalii factori de acțiune ai acestui sistem sunt 11 proteine, denumite C1-C9, B și D. Toate sunt prezente în mod normal printre proteinele plasmatice sanguine, precum și printre proteinele scurse din capilare în spațiile tisulare. Proenzimele nu sunt în mod normal active, dar pot fi activate în așa-numitul mod clasic. Complementul este principala componentă umorală a răspunsului imun înnăscut. La om, acest mecanism este activat prin legarea proteinelor complementului de carbohidrații de pe suprafața celulelor microbiene sau prin legarea complementului la anticorpii care s-au atașat acestor microbi. Un semnal sub forma unui complement atașat la membrana celulară declanșează reacții rapide care vizează distrugerea unei astfel de celule. Viteza acestor reacții se datorează creșterii rezultate din activarea proteolitică secvențială a moleculelor de complement, care sunt ele însele proteaze. Odată ce proteinele complement s-au atașat la un microorganism, acțiunea lor proteolitică este declanșată, care la rândul său activează alte proteaze ale sistemului complement și așa mai departe. Există trei căi de activare a complementului: clasică, lectină și alternativă. Lectina și căile alternative de activare a complementului sunt responsabile pentru reacția nespecifică a imunității înnăscute fără participarea anticorpilor. La vertebrate, complementul este, de asemenea, implicat în reacții specifice de imunitate, iar activarea lui are loc de obicei de-a lungul căii clasice. mod clasic Activarea complementului este un proces mediat imunologic inițiat de anticorpi. Specificitatea imunologică este asigurată de interacțiunea anticorpilor cu antigenele bacteriilor, virușilor și celulelor. Reacția antigen-anticorp este asociată cu o modificare a configurației imunoglobulinei, ceea ce duce la formarea unui situs de legare pentru Clq pe fragmentul Fc din apropierea regiunii balama. Imunoglobulinele se pot lega de C1. Activarea C1 are loc exclusiv între două fragmente Fc. Prin urmare, cascada de activare poate fi indusă chiar și de o singură moleculă de IgM. În cazul anticorpilor IgG, este necesară apropierea a două molecule de anticorpi, ceea ce impune restricții severe asupra densității epitopilor antigenului. În acest sens, IgM este un inițiator mult mai eficient al citolizei și al opsonizării imune decât IgG. Procesul de activare a complementului în sine poate fi împărțit în anumite etape: 1- recunoașterea complexelor imune și formarea C1; 2 - formarea C3-convertazei și C5-convertazei; 3 - formarea unui complex termostabil C5b, 6,7; 4 - perforarea membranei. Calea clasică este mai precisă, deoarece orice celulă străină este distrusă în acest fel. La cale alternativă anticorpii nu sunt implicați în activarea sistemului complementului. Principala diferență funcțională între reacția alternativă este viteza de răspuns la agentul patogen. În timp ce calea clasică de activare a complementului necesită timp pentru a acumula anticorpi specifici, o cale alternativă se dezvoltă imediat după intrarea agentului patogen. Inițiatorul procesului este C3b legat covalent de suprafața celulei. Secvența reacțiilor induse direct de microorganisme, care conduc la scindarea lui C3 și reglată de factorul I și factorul H se numește „calea alternativă a complementului”. Componenta C3 a complementului, prezentă abundent în plasmă, este împărțită constant în C3a și C3b. Legătura internă tioeterică din molecula nativă C3 este sensibilă la hidroliză spontană. Această activare spontană constantă, la nivel scăzut, a C3 plasmatic este denumită „martor” și menține o concentrație scăzută de C3b în plasmă. În ser, majoritatea C3b este inactivată prin hidroliză, dar unele C3b se leagă covalent la celulele gazdă sau la agenții patogeni invadatori. Legătura dintre C3b și agentul patogen este deosebit de semnificativă, deoarece contactul cu o suprafață străină determină un set de reacții care duc la acumularea ulterioară a C3b: într-o stare legată de celulă, C3b este capabil să interacționeze necovalent pe suprafață cu factorul. B. C3bB rezultat devine un substrat pentru proteaza serică - serin esteraza (factorul D). Factorul D desprinde un mic fragment de Ba din factorul B. Un fragment mare de Bb rămâne asociat cu C3b. Complexul C3bBb~ rezultat de pe suprafața agentului patogen se disociază foarte repede, cu excepția cazului în care este stabilizat prin legarea la properdin (factor P) și formând complexul C3bBbP~, care este o convertază C3 legată la suprafață de cale alternativă. Deoarece convertaza este localizată pe suprafața agentului patogen, moleculele C3b rezultate se vor lega acolo. Rezultatul unui lanț de reacții a căii alternative de activare a complementului este acumularea a doi factori de apărare nespecifici semnificativi: opsonina C3b și factorii inflamatori: C3a și C5b. Complexul C3bBb este stabilizat de properdin; în absența acesteia din urmă, complexul C3bBb este distrus rapid. Activarea căii alternative a complementului este inițiată de celule infectate cu anumiți virusuri, multe bacterii gram-pozitive și gram-negative, tripanozomi, leishmania, multe ciuperci, eritrocite heterologe, polizaharide, sulfat de dextran, precum și complexe de IgG, IgA sau IgE. cu antigen. Calea lectinei (manoză) de activare a sistemului complementului folosește lectina de legare a manozei (MBL), o proteină similară cu calea clasică de activare a C1q, care se leagă de reziduurile de manoză și alte zaharuri de pe membrană pentru a permite recunoașterea unei varietăți de agenți patogeni. MBL este o proteină serică aparținând grupului de proteine colectine, care este sintetizată în principal în ficat și poate activa cascada complementului prin legarea directă de suprafața agentului patogen. În serul sanguin, MBL formează un complex cu MASP-I și MASP-II (proteaza serină asociată cu lectina de legare a mananului, proteaze cu serin care leagă MBL). MASP-I și MASP-II sunt foarte asemănătoare cu C1r și C1 ale căii de activare clasice. Când mai multe situsuri active MBL se leagă într-un mod specific la reziduurile de manoză orientate de pe stratul dublu fosfolipidic al agentului patogen, MASP-I și MASP-II sunt activate și scindează proteina C4 în C4a și C4b și proteina C2 în C2a și C2b. C4b și C2a se combină apoi pe suprafața agentului patogen pentru a forma C3 convertaza, iar C4a și C2b acționează ca chemoatractanți pentru celulele sistemului imunitar.
Caracteristicile generale ale răspunsului imun la antigenele dependente de timus, etapele acestuia și rezultatul final.
De regulă, pentru a începe răspunsul imun (pentru majoritatea antigenelor), activarea T-helpers - Th. Antigenele, răspunsul la care se dezvoltă cu ajutorul lui Th, se numesc dependenți de timus, iar răspunsul în sine se numește răspuns imun dependent de timus.
Antigenele dependente de timus se numesc antigene, formarea de anticorpi împotriva cărora necesită o cooperare complexă a macrofagelor, limfocitelor T și B.
Răspunsul imun la aceste antigene este caracterizat prin următoarele etape.
4) transferul de informații despre antigen către un al treilea grup de celule imunocompetente (fie către macrofage specializate - așa-numitul tip celular de răspuns imun implementat de subtipul 1 T-helper, fie către limfocite B - un tip de răspuns imun care conduce la producerea de anticorpi specifici antigenului care a provocat răspunsul imun și implementați de subtipul T-helper 2);
Dezvoltarea și caracteristicile celulelor prezentatoare de antigen, localizarea lor în organism
Celule prezentatoare de antigen (APC) - captează antigenele, le procesează și prezintă determinanții antigenici corespunzători altor celule imunocompetente. Există două tipuri de celule prezentatoare de antigen: „profesionale” și „non-profesionale”. "Profesional" celulele prezentatoare de antigen captează antigenul foarte eficient prin fagocitoză sau endocitoză mediată de receptor și apoi prezintă un fragment din acest antigen pe membrana lor în complex cu moleculele MHC clasa II. Celulele T recunosc acest complex pe membrană și interacționează cu acesta. Celulele prezentatoare de antigen produc apoi molecule co-stimulatoare suplimentare, rezultând activarea celulelor T. Expresia acestor molecule co-stimulatoare este o trăsătură caracteristică celulelor „profesionale” prezentatoare de antigen. Există mai multe tipuri principale de celule „profesionale” prezentatoare de antigen: celulele dendritice , care sunt cele mai importante celule prezentatoare de antigen. Celulele dendritice activate sunt activatori T-helper deosebit de eficienți, deoarece moleculele costimulatoare, cum ar fi proteina B7, sunt prezente pe suprafața lor. macrofage , care sunt celule CD4-pozitive și, prin urmare, pot fi infectate cu virusul imunodeficienței umane. limfocitele B , care poartă pe suprafața lor (ca receptor de celule B) și secretă anticorpi specifici și pot captura, de asemenea, antigenul legat de receptorul de celule B, îl prelucrează și îl prezintă într-un complex cu molecule de complex major de histocompatibilitate de clasa II. În raport cu alte tipuri de antigene, limfocitele B sunt inactive ca celule prezentatoare de antigen. Unele celule epiteliale activate. Celulele dendritice, precum macrofagele și limfocitele, sunt de origine hematopoietică. Celulele dendritice sunt localizate în epiteliul intestinal, tractul urogenital, căile respiratorii, plămâni, epiderma pielii (celule Langerhans) și spațiile interstițiale. „Neprofesionist » Celulele prezentatoare de antigen nu conțin în mod normal molecule de clasa II ale complexului major de histocompatibilitate, ci le sintetizează numai ca răspuns la stimularea cu anumite citokine, de exemplu, interferonul y. Celulele non-profesionale prezentatoare de antigen includ:
fibroblaste cutanate
celule epiteliale timusului
celulele epiteliale tiroidiene
celule gliale
celulele β ale pancreasului
celule endoteliale vasculare
Celulele prezentatoare de antigen sunt prezente predominant în piele, ganglioni limfatici, splină și timus.
Procesarea antigenului, importanța sa în dezvoltarea răspunsului imun
procesarea antigenelor. Expresia moleculelor HLA din clasele I și II care prezintă un antigen este reglată de trei loci genetici HLA - TAP, DM și LMP, care determină interacțiunea acestora cu antigenele. Moleculele HLA-LMP 2 și HLA-LMP 7, care sunt exprimate sub influența interferonului gamma, sunt primele incluse în sistemul de procesare a diferitelor antigene exogene. Acestea declanșează proteoliza în proteazomi și reglează dimensiunea și specificitatea peptidelor pentru legarea la moleculele HLA. Proteazomul este un complex enzimatic format din 24 de subunități proteice. Două lanțuri de molecule HLA clasa II sunt sintetizate în reticulul endoplasmatic, conectate temporar la al treilea lanț, invariant II (CD74), care împiedică legarea lor de autopeptide. Apoi, acest complex este transferat la endozomi, unde se leagă de peptida antigen corespunzătoare cu lungimea de 9-25 aminoacizi, care înlocuiește lanțul II invariant. Prin fuziunea endozomului cu membrana, moleculele HLA-DR sunt exprimate cu un antigen-peptid pe suprafata celulei. Deplasarea peptidei lanțului invariant și înlocuirea acesteia cu o peptidă antigen specifică sunt efectuate de proteine speciale ale locusului HLA-DM care catalizează acest proces. Moleculele MHC clasa I sunt sintetizate constant în reticulul endoplasmatic al celulei și stabilizate de proteina calnexina. Antigenii endogeni și virali sunt preclivați în proteazom în peptide cu 8-11 resturi de aminoacizi în dimensiune. Când se leagă de antigen-peptidă, kalnexina este scindată, iar moleculele MHC sunt transferate folosind proteinele de transport HLA-TAP (transportor of antigen processing) la suprafața celulei, unde acest complex este prezentat supresoarelor T/ucigașilor. Caracteristicile structurale ale moleculelor MHC clasa II, spre deosebire de MHC clasa I, sunt de așa natură încât asigură legarea mai multor peptide antigene polimorfe. Moleculele MHC capătă o formă tridimensională stabilă pe celule numai după ce sunt legate de situsurile de pliuri ale peptidelor corespunzătoare. Complexul „MHC-peptidă moleculă” prezentat rămâne pe celulă (macrofag, etc.) timp de câteva săptămâni, ceea ce permite altor celule, în special limfocitelor T, să interacționeze cu aceasta. Specificitățile alelice specifice ale moleculelor MHC intră în legătură cu un peptid-antigen specific, care asigură recunoașterea antigenului. De exemplu, o peptidă de virus herpes se leagă de haplotipul HLA-DQA 1*0501/DQB 1*2001, dar nu de altul care diferă doar prin 15 reziduuri de aminoacizi.
Limfocitele T, dezvoltarea și localizarea lor. T-helper și rolul lor în dezvoltarea răspunsului imun la antigenele dependente de timus
Timusul oferă condiții optime pentru dezvoltarea tuturor subpopulațiilor de limfocite T de la precursorii măduvei osoase la formele mature cu TCR cu drepturi depline. Celulele epiteliale joacă un rol cheie în micromediul limfocitelor T din timus. Ei sunt cei care asigură condițiile necesare pentru diferențierea limfocitelor T. Focarele de dezvoltare extra-timică a celulelor T existente în organism (de exemplu, în intestin) nu oferă un astfel de efect în totalitate. Odată cu vârsta, fondul de limfocite T naive care părăsesc timusul scade. În acest moment, sistemul imunitar „folosește” celulele T de memorie formate în organism. Una dintre problemele cheie ale sistemului imunitar adaptativ la vârstnici este capacitatea de a răspunde în mod adecvat la noi antigene pe care organismul nu i-a mai întâlnit înainte (de exemplu, „noile” boli infecțioase sunt mai severe decât la o vârstă fragedă și mai des. duce la complicații și deces) . Principalele etape ale dezvoltării limfocitelor T în timus (imunopoieza celulelor T) au fost determinate în conformitate cu un program determinat genetic și în absența stimulării antigenice: formarea receptorilor de recunoaștere a antigenului specific clonelor capabili să recunoască peptidele antigenice în combinație cu molecule HLA autologe; eliminarea celulelor T specifice auto-antigenelor; expresia moleculelor co-receptoare CD4 sau CD8 cu formarea subpopulațiilor de T-helper și CTL, precum și a celulelor T reglatoare naturale (naturale) (Treg). Diferențierea în timus este însoțită de o modificare a markerilor de suprafață ai T. -limfocite. Include următoarele etape: migrarea precursorilor celulelor T din măduva osoasă; rearanjarea genelor TCR și formarea unui receptor complet; selecția pozitivă și negativă a celulelor T; formarea de subpopulații mature de limfocite T CD4+ și CD8+; emigrarea celulelor T mature din timus. Progenitorii limfoizi timpurii (CD34, CD38, CD45RA, CD117, CD7, CD44) formați în ficatul fetal și ulterior în măduva osoasă intră în parenchimul timusului prin diapedeză prin venule postcapilare cu endoteliu ridicat situat la joncțiunea cortico-medulară și se deplasează. spre straturile exterioare ale cortexului și apoi migrează din nou în zona joncțiunii corticomedulare. Când celulele migrează, ele se diferențiază.
Dacă la o suspensie celulară constând din macrofage, limfocite T și limfocite B a fost adăugat un antigen capabil să provoace proliferarea (o creștere a numărului) celulelor B, s-a observat ca rezultat un răspuns proliferativ bine definit de la celulele B. Dacă suspensia celulară a constat numai din limfocite T și B, răspunsul proliferativ al acestora din urmă nu a fost înregistrat. Dacă antigenul a fost adăugat într-o suspensie constând numai din macrofage, păstrată o perioadă de timp, iar apoi, după eliberarea suspensiei de excesul de antigen, a fost amestecată cu limfocite T și B, s-a observat o proliferare pronunțată a celulelor B.
Studii mai detaliate ale rolului macrofagelor în aceste procese nu numai că au confirmat rolul lor inițiator, dar au făcut și posibilă descrierea mecanismului participării lor la formarea răspunsului imun. Informațiile obținute în acest fel au stat la baza schemei acum general acceptate de interacțiune celulară cu trei cooperare în timpul dezvoltării unui răspuns imun la antigenele dependente de timus.
Conform acestei scheme, în organismul care răspunde la pătrunderea antigenului, se întâmplă următoarele:
1) perceperea și prelucrarea informațiilor conținute în antigen de către celulele sistemului macrofag;
2) transmiterea acestor informații către celulele sistemului limfocitar și anume T-limfocite-helpers (T-helpers);
3) activarea T-helpers care au primit informația și proliferarea acestora;
4) transferul de informații despre antigen către un al treilea grup de celule imunocompetente (fie către macrofage specializate - așa-numitul tip celular de răspuns imun implementat de subtipul 1 T-helper, fie către limfocite B - un tip de răspuns imun care conduce la producerea de anticorpi specifici antigenului care a provocat răspunsul imun și implementați de subtipul T-helper 2);
5) activarea celulelor celui de-al treilea grup care au primit informația și fie distrugerea de către macrofage activate a propriilor celule modificate prin acțiunea antigenului (răspuns imun de tip celular), fie formarea de către limfocite B activate a unei multitudini. de anticorpi care interacționează în mod specific cu antigenul care a provocat răspunsul imun (răspunsul imun de tip producator de anticorpi).
Limfocitele B, dezvoltarea și localizarea lor. Producția de celule plasmatice și anticorpi
Dezvoltarea limfocitelor B pe parcursul întregii perioade postembrionare are loc în măduva osoasă. Sub influența micromediului celular al măduvei osoase și a factorilor umorali ai măduvei osoase, din celula stem limfoidă se formează limfocitele B. Stadiile incipiente ale dezvoltării limfocitelor B depind de interacțiunea de contact direct cu elementele stromale. Etapele ulterioare ale dezvoltării limfocitelor B au loc sub influența factorilor umorali ai măduvei osoase. Interacțiunea celor mai timpurii precursori ai celulelor B (limfocite pro B timpurii) cu elementele stromale se realizează cu ajutorul moleculelor adezive de suprafață CD44, c-kit și SCF. Ca urmare a acestor contacte, are loc o creștere a proliferării limfocitelor B și trecerea lor la următoarea etapă de dezvoltare - celulele pro-B tardive. Receptorul IL-7 este exprimat pe suprafața celulelor pro-B tardive. Sub influența IL-7 produsă de elementele stromale, limfocitele pro-B proliferează și se diferențiază în celule pre-B timpurii, caracterizate prin prezența în citoplasma lor a lanțului m-polipeptidic de imunoglobuline. Aceste celule au morfologia celulelor limfoide mari. Ulterior, aceste celule se transformă în limfocite pre-B mici, în unele dintre ele, pe lângă lanțul polipeptidic m-greu, în citoplasmă sunt detectate lanțuri ușoare de imunoglobuline. În următoarea etapă de dezvoltare a limfocitelor B, are loc expresia imunoglobulinelor monomerice de suprafață M. Aceste structuri sunt receptorii care recunosc antigenul ai celulelor B. Specificitatea antigenică a receptorilor este determinată genetic. La următoarea etapă de dezvoltare a limfocitelor B, celulele sunt orientate spre sinteza anticorpilor dintr-o anumită clasă. Apar limfocitele B care, împreună cu IgM, exprimă molecule din clasa IgA sau IgG. Aceasta este urmată de exprimarea pe celulele IgD. Odată cu exprimarea imunoglobulinelor D pe limfocite, etapa de maturare independentă de antigen a celulelor B este finalizată. Astfel, pe limfocitele B mature, moleculele de Ig de suprafaţă pot fi reprezentate prin următoarele clase: 1) IgM, IgD; 2) IgM, IgA, IgD; 3) IgM, IgG, IgD. Mai mult, toate imunoglobulinele prezente pe o celulă B au același idiotip, deoarece sunt codificate de aceleași gene VH și VL. Exprimarea moleculelor MHC pe limfocitele B se observă pornind de la stadiul pro-celulă B. Aceste antigene sunt exprimate pe toate celulele B mature. Receptorii pentru componenta C3 a complementului (RC3b) și fragmentul Fc al Ig (RFc) sunt detectați mai întâi în cantități mici pe celulele B imature. Pe celulele mature, aceste molecule au o densitate mare și sunt ușor de detectat. Limfocitele B mature se caracterizează prin prezența IgD de suprafață, o densitate mare a receptorilor pentru componenta complementului C3 și fragmentul Fc al Ig, capacitatea de a se transforma în forme blastice sub influența mitogenilor B (LPS, PWM), și capacitatea de a se transforma sub influența antigenilor în celule formatoare de anticorpi.
memorie imunologică. Răspunsul imun primar și secundar
memorie imunologică este capacitatea sistemului imunitar de a răspunde mai rapid și mai eficient la un antigen (patogen) cu care organismul a avut contact anterior.
Sistemul imunitar are două proprietăți cu adevărat uimitoare: recunoașterea specifică și memoria imunitară. Acesta din urmă este înțeles ca capacitatea de a dezvolta un răspuns imun mai eficient calitativ și cantitativ la contactul repetat cu același agent patogen. În consecință, se face o distincție între răspunsurile imune primare și secundare. Răspunsul imun primar se realizează la primul contact cu un antigen necunoscut, iar cel secundar la contactul repetat. Răspunsul imun secundar este mai perfect, deoarece este realizat la un nivel calitativ mai ridicat datorită prezenței unor factori imunitari preformați care reflectă adaptarea genetică la agentul patogen (există deja gene gata făcute pentru imunoglobuline specifice și recunoașterea antigenului T-). receptorii celulari). Într-adevăr, oamenii sănătoși nu se îmbolnăvesc de două ori de multe boli infecțioase, deoarece atunci când sunt reinfectate, se realizează un răspuns imun secundar, în care nu există o fază inflamatorie pe termen lung, iar factorii imuni - limfocite și anticorpi specifici - imediat intră în joc.
Răspunsul imun secundar este caracterizat de următoarele caracteristici:
unu . O dezvoltare mai timpurie, uneori chiar fulgerătoare.
2. Este necesară o doză mai mică de antigen pentru a obține un răspuns imun optim.
3 . O creștere a puterii și a duratei răspunsului imun datorită producției mai intense de citokine (profiluri TD 1 sau 2, în funcție de natura agentului patogen).
4 . Întărirea răspunsurilor imune celulare datorită formării mai intense a T-helper specific de tip 1 și a limfocitelor T citotoxice.
5 . Creșterea formării de anticorpi datorită formării mai multor ajutoare T - tip 2 și celule plasmatice.
6. O creștere a specificității recunoașterii peptidelor imunogene de către limfocitele T datorită creșterii afinității receptorilor lor specifici pentru antigen.
7. O creștere a specificității anticorpilor sintetizați datorită producției inițiale de IgG de mare afinitate/aviditate.
Trebuie remarcat faptul că imposibilitatea formării unei memorii imunitare eficace este unul dintre simptomele caracteristice bolilor imunodeficienței umane. Deci, la pacienții cu hipoimunoglobulinemie, fenomenul de episoade multiple de așa-numita. infecții din copilărie, deoarece după boli infecțioase nu se formează un titru de anticorpi protector. Pacienții cu defecte ale imunității celulare, de asemenea, nu formează o memorie imună pentru antigenele dependente de T, care se manifestă prin absența seroconversiei după infecții și vaccinări, cu toate acestea, concentrațiile totale de imunoglobuline din serul lor sanguin pot fi normale.
Natura interacțiunilor celulelor prezentatoare de antigen, limfocitelor T și B în timpul dezvoltării unui răspuns imun la antigenele dependente de timus, rolul antigenilor de suprafață (proteinele complexului major de histocompatibilitate și altele) în aceste interacțiuni
Celulele prezentatoare de antigen sunt prezente predominant în piele, ganglioni limfatici, splină și timus. Acestea includ macrofage, celule dendritice, celule proces foliculare ale ganglionilor limfatici și ale splinei, celule Langerhans, celule M din foliculii limfatici ai tractului digestiv, celule epiteliale ale glandei timus. Aceste celule captează, procesează și prezintă Ag (epitop) pe suprafața lor altor celule imunocompetente, produc citokine, secretă prostaglandina E2, care suprimă răspunsul imun. Celulele dendritice provin din măduva osoasă și formează o populație de celule cu viață lungă care declanșează și modulează răspunsul imun. În măduva osoasă, progenitorii lor formează o subpopulație de celule CD34+ care sunt capabile să se diferențieze în celule Langerhans pentru epiteliu și celule dendritice pentru mediul intern. Precursorii celulelor dendritice imature și nedivizabile colonizează multe țesuturi și organe. Celulele dendritice sunt stelate și poartă un număr relativ mic de molecule MHC pe suprafața lor în repaus. Spre deosebire de celulele Langerhans, celulele dendritice interstițiale sunt capabile să stimuleze sinteza Ig de către limfocitele B. Varietăți de DC: - mieloide - provin din monocite. Ele pot fi considerate ca un fel de macrofage specializate în prezentarea Ag la limfocitele T; - limfoizii provin dintr-o celulă progenitoare limfoidă comună, din care se dezvoltă și limfocitele T și B. Interacțiunea limfocitelor T și B.În răspunsul imun primar, singurele APC eficiente pentru limfocitele T sunt DC. Dar în cazul activării limfocitelor T de către Ag reprezentat de DC, limfocitele B adiacente vor fi și ele implicate în răspunsul imun. În acest caz, sunt posibile două opțiuni pentru interacțiunea limfocitelor T și B:
Limfocitele B leagă Ag solubil cu receptorul lor de imunoglobulină, îl absorb prin endocitoză, îl procesează în interiorul lor și expun fragmente de Ag la suprafață ca parte a complexelor cu moleculele MHC-II și MHC-I. TCR-ul limfocitului T leagă Ag pe suprafața limfocitului B, acționând ca APC; în plus, sunt stabilite toate relațiile coreceptoare necesare și suficiente între limfocitele T și B. O astfel de interacțiune are loc în zonele dependente de T ale țesutului limfoid periferic la începutul dezvoltării răspunsului imun.
Un limfocit B își recunoaște Ag, dar un limfocit T care recunoaște Ag pe un alt APC și este activat prin interacțiunea cu acest alt APC nu va fi departe. În acest caz, interacțiunea T-B poate fi limitată de interacțiunea citokinelor limfocitelor T cu receptorul acestor citokine de pe limfocitul B, iar interacțiunea moleculelor de membrană între ele poate sau nu să apară într-o oarecare măsură (cel puțin în răspunsul imun primar). ). Dar în răspunsul imun secundar, interacțiunea moleculei de membrană a limfocitului B CD40 cu molecula de membrană a limfocitului T CD40L are loc în mod necesar, deoarece fără această interacțiune nu există nicio schimbare a clasei de imunoglobuline de la IgM la altele, iar răspunsul secundar al limfocitelor B2 se caracterizează prin comutarea obligatorie a clasei de imunoglobuline cu IgM pentru IgG, IgA sau IgE. Aceste interacțiuni T-B apar deja pe teritoriul zonelor celulelor B - în foliculii organelor limfoide. Antigenii complexului major de histocompatibilitate (MHC) sunt un grup de proteine de suprafață ale diferitelor celule ale corpului care joacă un rol cheie în răspunsurile imune mediate de celule. Moleculele codificate de MHC se leagă de antigenele peptidice, drept urmare acești antigeni sunt recunoscuți de receptorii specifici de pe limfocitele T și B. Limfocitele T citotoxice (T-killers) recunosc celulele țintă numai dacă au antigene MHC clasa I cu propriul lor genotip pe suprafața lor. Când celulele care interacționează în răspunsul imun poartă diferite alele MHC, răspunsul imun se dezvoltă nu împotriva antigenului străin prezentat (de exemplu, viral sau bacterian), ci împotriva diferiților antigeni MHC. Acest fenomen stă la baza faptului că antigenele MHC asigură recunoașterea „sinelui” și a „străinilor” în organism.
Conceptul de antigene. Proprietățile generale ale antigenelor. Antigene complete și incomplete.
Antigenele sunt numite substanțe străine din punct de vedere structural pentru acest organism special (compuși cu molecul mare - proteine și polizaharide) care pot provoca un răspuns imun.
Principalele proprietăți ale antigenelor:
- străinătate. Conceptul de antigene nu poate fi separat de conceptul de străinătate. Folosim termenul de antigen, adică străinătatea lui în raport cu un organism dat. De exemplu, pentru o persoană, proteina unui animal sau a altei persoane va fi un antigen.
Strainătatea este determinată de greutatea moleculară, dimensiunea și structura biopolimerului, rigiditatea macromoleculară și structurală a acestuia.
- antigenicitate. Antigenicitatea proteinelor este o manifestare a străinătății lor, iar specificitatea acesteia depinde de secvența de aminoacizi a proteinelor, structura secundară, terțiară și cuaternară (adică de conformația globală a moleculei proteice), de grupele determinante localizate superficial și de aminoacizi terminali. reziduuri acide. Starea coloidală și solubilitatea sunt proprietăți esențiale ale antigenelor.
- imunogenitate. Imunogenitatea este capacitatea de a induce un răspuns imun cu formarea de anticorpi, adică de a forma imunitate. Conceptul de imunogenitate se referă în principal la antigenele microbiene care asigură formarea imunității, adică imunitate la infecții.
Imunogenitatea depinde de o serie de factori (greutatea moleculară, mobilitatea moleculelor de antigen, formă, structură, capacitatea de schimbare).
- specificitatea. Conceptul de specificitate antigenă se referă la caracteristicile care disting un antigen de altul.
Antigenul ca principală cauză a dezvoltării procesului imunitar a fost de interes pentru imunologi încă de la începuturile imunologiei. Cu toate acestea, doar datorită cercetărilor lui Karl Landsteiner din anii 1920 și 1930 au fost create condițiile pentru studierea naturii subtile a specificității antigenului. Compușii organici simpli au fost luați ca material antigenic - haptene . Prin ei înșiși, acești compuși nu sunt capabili să provoace o reacție imunologică. Prezența străinilor la greutate moleculară mică îi privează de imunogenitate. Complexul haptenei cu proteina purtătoare este imunogen.
În caz contrar, haptenele sunt cunoscute ca antigene incomplete.. De regulă, au o greutate moleculară mică și nu sunt recunoscute de celulele imunocompetente. Haptenele pot fi simple sau complexe; haptenele simple interacționează cu anticorpii din organism, dar nu sunt capabile să reacționeze cu aceștia in vitro; haptenele complexe interacționează cu anticorpii in vivo și in vitro. Haptenele pot deveni imunogene atunci când sunt legate de un purtător cu greutate moleculară mare care are propria sa imunogenitate.
În funcție de origine, antigenele sunt clasificate în exogene, endogene și autoantigene.
exogene antigenii pătrund în organism din mediul înconjurător, prin inhalare, ingerare sau injectare. Astfel de antigeni intră în celulele prezentatoare de antigen prin endocitoză sau fagocitoză și apoi sunt procesați în fragmente. Celulele prezentatoare de antigen prezintă apoi fragmente la celulele T-helper (CD4+) pe suprafața lor prin intermediul moleculelor de complex major de histocompatibilitate de tip II (MHC II).
Endogen antigenele sunt produse de celulele corpului în timpul metabolismului natural sau ca urmare a unei infecții virale sau bacteriene intracelulare. Fragmentele sunt apoi prezentate pe suprafața celulei într-un complex cu proteine ale complexului major de histocompatibilitate tip 1 MHC I. Dacă antigenele prezentate sunt recunoscuți de limfocitele citotoxice, celulele T secretă diverse toxine care provoacă apoptoza sau liza celulei infectate. Pentru a preveni limfocitele citotoxice să distrugă celulele sănătoase, limfocitele T autoreactive sunt excluse din repertoriu în timpul selecției pentru toleranță.
Autoantigene sunt proteine normale sau complexe proteice care sunt recunoscute de sistemul imunitar la pacientii cu boli autoimune. Astfel de antigeni nu ar trebui să fie în mod normal recunoscuți de sistemul imunitar, dar din cauza factorilor genetici sau de mediu, toleranța imunologică la astfel de antigene se poate pierde la acești pacienți.
Tipuri de specificitate antigenică.
1) specificitatea speciei- se referă la prezența antigenelor caracteristice tuturor indivizilor unei specii și necaracteristice organismelor altor specii;
2) heterospecificitate- specificitatea antigenică, datorită prezenței antigenelor comune pentru reprezentanții de diferite tipuri.
3) specificitatea grupului- diferențe de antigene ale grupurilor de indivizi din cadrul unei specii, de exemplu, împărțirea oamenilor în funcție de antigenele eritrocitare în așa-numitele grupe sanguine;
4) specificitatea tipului- un concept care practic coincide cu specificul de grup, dar este folosit pentru speciile microbiene;
5) specificitatea funcțională- asemănarea în determinanții antigenici ai moleculelor care îndeplinesc aceeași funcție în diferite organisme. astfel de molecule au nu numai determinanți similari, ci și cei prin care se manifestă specificitatea speciei sau a grupului, datorită cărora este posibil să se distingă, de exemplu, enzime cu aceeași specificitate de substrat formate în organisme ale animalelor de specii diferite;
6) specificitatea etapei- un concept legat de embriogeneză: vorbim de molecule care apar doar într-un anumit stadiu al dezvoltării embrionare și sunt absente, în alte stadii de ontogeneză. Detectarea unor astfel de antigene face posibilă determinarea stadiului de dezvoltare cu mare precizie, mai ales când diferențierea morfologică și anatomică a stadiilor este dificilă sau imposibilă;
7) specificitatea patologică- prezența antigenelor care sunt necaracteristice pentru organism în normă și apar numai în patologie. detectarea lor deschide noi posibilități pentru diagnosticarea unui număr de boli (de exemplu, modificări maligne) și monitorizarea stării pacienților în timpul terapiei;
8) specificitatea haptenelor- proprietăţi ale antigenelor complexe determinate de o haptenă specifică. Este important în dezvoltarea răspunsurilor imune la substanțele cu greutate moleculară mică, în special la antibiotice sau coloranți cu anilină, la care persoanele cu anumite profesii pot fi alergice. Pentru a detecta specificitatea antigenică a oricăruia dintre aceste tipuri, se utilizează suspensii adecvate de anticorpi sau imunoglobuline.
Haptene- Sunt antigene de natura organica inrudite cu lipide si polizaharide.
Dependența proprietăților antigenice de structura moleculară.
Antigenicitatea unei molecule este determinată de capacitatea sa de a induce un răspuns imun într-un anumit organism. Antigenicitatea implică capacitatea moleculelor de a fi recunoscute individual de receptorii celulelor imunocompetente, adică. această proprietate determină specificitatea răspunsului imun. Majoritatea antigenelor (în principal de natură proteică) pot determina formarea memoriei imunologice. Acest lucru este important în legătură cu antigenele microorganismelor care provoacă imunitate la infecție - cât de imunogen este acest sau acel vaccin.
Gradul de antigenicitate depinde de o serie de factori. De mare importanță este dimensiunea și greutatea moleculară a antigenului. Cu cât greutatea moleculară a moleculei este mai mare, cu atât sunt mai puternice proprietățile sale antigenice.
Determinant antigenic
Determinant antigenic [gr. anti - contra și gene - generative; lat. determinantis - limitator, definitor] - partea structurală a antigenului de care se leagă anticorpul. Iad. constă din mai mulți aminoacizi (de obicei 6-8), formând o structură spațială caracteristică unei anumite proteine. Într-o proteină, constând din câteva sute de aminoacizi, există mai multe (5-15) diferite A.D. Au fost dezvoltate programe speciale pentru a prezice localizarea proteinei A.d., recunoscută în timpul răspunsului imun umoral, ceea ce face posibilă utilizarea pentru imunizare nu a proteinelor întregi, ci a peptidelor scurte care conțin A.d.
Determinanții pot fi extrem de diverși în formă și distribuție a sarcinilor și contribuie la dezvoltarea unor răspunsuri destul de diverse ale răspunsului imun umoral.
Antigene: valență
Valenta antigenului este numărul de situsuri de legare a anticorpilor. Această valoare poate varia semnificativ în funcție de structura antigenului, de mărimea acestuia, precum și de tipul de animal de la care au fost obținuți anticorpii.
Antigenele, de regulă, poartă mulți factori determinanți. Cu cât molecula de antigen este mai mare, cu atât determinantul conține mai mult, cu atât este mai mare valența sa. Antigenele poartă de obicei determinanți cu specificitate diferită. Ca rezultat, introducerea majorității antigenelor are ca rezultat formarea de anticorpi cu specificitate diferită.
Clasificarea antigenelor după origine. Tipuri de specificitate antigenică
Anticorpii sunt gammaglobuline specifice serului sanguin, formate ca răspuns la introducerea de antigene sau ca urmare a contactului natural al organismului cu substanțe antigenice (bacterii, toxine, proteine de diverse origini, polizaharide, complexe polizaharid-proteine etc.). Pentru a produce o cantitate semnificativă de anticorpi, o cantitate mică de antigen intră în organism. Unitatea structurală de bază (monomer) a unei imunoglobuline de orice clasă constă din două lanțuri polipeptidice ușoare identice (L - din engleză light) și două lanțuri polipeptidice grele identice (H - din engleză heavy) ținute împreună prin legături disulfurice. Lanțurile ușoare conțin 2 regiuni omoloage, iar lanțurile grele, în funcție de clasa de imunoglobuline, 4-5 regiuni omoloage, formate din aproximativ 110 resturi de aminoacizi și având structuri globulare ținute împreună printr-o legătură disulfură și având funcții autonome. Astfel de structuri se numesc domenii. Centrii de legare a antigenului ai imunoglobulinelor sunt formați din secvențele N-terminale ale lanțurilor ușoare și grele, adică domenii variabile ale acestor lanțuri (domenii V). Mai multe (3-4) regiuni hipervariabile sunt izolate în domeniile V. Structura domeniilor rămase este constantă, deci sunt numite constante sau domenii C. Lanțurile ușoare conțin un domeniu C, lanțurile grele conțin 3-4 domenii C. Sub influența papainei, moleculele de imunoglobulină (monomeri) sunt scindate cu formarea a două fragmente Fab (legare la antigen fragment) care leagă antigenul și un fragment Fc (fragment cristalizabil, constant), care este partea C-terminală a moleculă, formând ușor cristale. Fragmentele Fc din cadrul aceleiași clase sunt identice (constante), indiferent de specificitatea imunoglobulinelor. Acestea asigură interacțiunea complexelor antigen-anticorp cu proteinele complementului, fagocite, eozinofile, bazofile și mastocite. Moleculele IgG, IgD și IgE sunt monomeri, IgM - pentameri; Moleculele de IgA din sânge sunt monomeri, în saliva și secrețiile mucoasei sunt dimeri. Imunoglobulina M (lgM) se formează într-un stadiu incipient al răspunsului imun și indică un proces infecțios acut. În molecula IgM, cinci subunități sunt conectate printr-un lanț J (din engleză joining - binding), ca urmare a căruia molecula are 10 centri de legare a antigenului.
Imunoglobulina A (lgA) se găsește pe suprafața mucoaselor, în colostru, lapte, salivă și lichid lacrimal. Conține o componentă secretorie care este sintetizată în celulele epiteliale și o protejează de scindarea de către enzimele proteolitice. Imunoglobulina E (lgE) are forma unei subunități monomer (L-H) 2 și o greutate moleculară de aproximativ 190 000. Este conținută în urme în serul sanguin. Are o activitate homocitotropă ridicată, adică se leagă puternic de mastocite ale țesutului conjunctiv și de bazofilele din sânge. Interacțiunea IgE legată de celule cu un antigen înrudit provoacă degranularea mastocitelor, eliberarea de histamină și alte substanțe vasoactive, ceea ce duce la dezvoltarea unei hipersensibilități imediate. Anterior, anticorpii din clasa IgE erau numiți reagine. Imunoglobulina D (lgD) există ca anticorp monomeric cu o greutate moleculară de aproximativ 180 000. Concentrația sa în serul sanguin este de 0,03-0,04 g/l. lgD este prezent ca receptor pe suprafața limfocitelor B.
FuncțiiFAb- șiFc-părți ale unei molecule de imunoglobulină
Moleculele de imunoglobuline din toate cele cinci clase constau din lanțuri polipeptidice: două lanțuri grele identice H și două lanțuri uşoare identice - L, conectate prin punți disulfurice. Conform fiecărei clase de imunoglobuline, i.e. M, G, A, E, D disting cinci tipuri de lanțuri grele: μ (mu), γ (gamma), α (alfa), ε (epsilon) și Δ (delta), care diferă în antigenicitate. Lanțurile ușoare din toate cele cinci clase sunt comune și vin în două tipuri: κ (kappa) și λ (lambda); Lanțurile L ale imunoglobulinelor din diferite clase se pot uni (recombina) atât cu lanțuri H omoloage, cât și cu cele heterologe. Cu toate acestea, numai lanțurile L identice (κ sau λ) pot fi în aceeași moleculă. Ambele lanțuri H și L au o regiune variabilă - V, în care secvența de aminoacizi este instabilă și o regiune constantă - C cu un set constant de aminoacizi. În lanțurile ușoare și grele, se disting grupările terminale NH2 și COOH.
La procesarea γ-globulinei mercaptoetanol legăturile disulfurice sunt distruse și molecula de imunoglobulină se rupe în lanțuri individuale de polipeptide. Când este expus la o enzimă proteolitică papaină imunoglobulina este scindată în trei fragmente: două fragmente necristalizante care conţin grupări determinante pentru antigen şi numite fragmente Fab I şi II, şi un fragment Fc cristalizant. Fragmentele FabI și FabII sunt similare ca proprietăți și compoziție de aminoacizi și diferă de fragmentul Fc; Fragmentele Fab și Fc sunt formațiuni compacte interconectate prin secțiuni flexibile ale lanțului H, datorită cărora moleculele de imunoglobulină au o structură flexibilă.
Papaina împarte molecula de imunoglobulină în două identice Fab - Fragment (Legarea antigenului fragmentului), fiecare dintre ele având un centru de legare a antigenului și Fc-fragment(Fragment cristalizabil) incapabil de a lega antigenul.
Pepsina scindează molecula în altă parte, tăind fragmentul pFc" din fragmentul mare 5S, numit F(ab") 2 deoarece, ca și anticorpul părinte, este bivalent pentru legarea antigenului. Fragmentul pFc" reprezintă partea C-terminală a regiunii Fc, regiunea lanţului greu din fragmentul Fab este desemnată Fd.
Studiile au arătat că o parte a anticorpului (fragmentul Fab) este concepută pentru a se lega de antigen, iar cealaltă parte (fragmentul Fc) interacționează cu celulele sistemului imunitar: neutrofile, macrofage și alte fagocite mononucleare care poartă receptori pentru Fc. fragment pe suprafața lor. Prin urmare, dacă anticorpii se leagă de microorganisme patogene, ei pot interacționa și cu fagocitele cu fragmentul lor Fc. Din acest motiv, celulele agentului patogen vor fi distruse de aceste fagocite. De fapt, anticorpii acționează în acest caz ca molecule intermediare.
27.
Clasele de imunoglobuline de mamifere. Diferențele structurale și funcționale ale imunoglobulinelor din diferite clase
(IgG) alcătuiesc aproximativ 80% din imunoglobulinele serice, cu un mol. cântărind 160 000. Ele se formează la înălțimea răspunsului imun primar și la administrarea repetată a antigenului (răspunsul secundar). IgG-urile au o rată mare de legare a antigenului, în special cele de natură bacteriană. Aceasta determină capacitatea IgG de a participa la reacțiile de protecție ale bacteriolizei. IgG este singura clasă de anticorpi care traversează placenta în făt. La ceva timp după nașterea unui copil, conținutul său în serul sanguin scade și atinge o concentrație minimă cu 3-4 luni, după care începe să crească din cauza acumulării propriilor IgG, ajungând la norma până la vârsta de 7 ani. . Dintre toate clasele de imunoglobuline, IgG este cea mai sintetizată în organism. Aproximativ 48% din IgG este conținută în lichidul tisular în care difuzează din sânge. IgG, precum și imunoglobulinele din alte clase, suferă o degradare catabolică, care are loc în ficat, macrofage și focar inflamator sub acțiunea proteinazelor.
(IgM) sunt primele care sunt sintetizate în corpul fătului și primele care apar în serul sanguin după imunizarea persoanelor cu majoritatea antigenelor. Ele reprezintă aproximativ 13% din imunoglobulinele serice la o concentrație medie de 1 g/l. În ceea ce privește greutatea moleculară, acestea sunt semnificativ superioare tuturor celorlalte clase de imunoglobuline. Acest lucru se datorează faptului că IgM sunt pentameri, adică sunt formați din 5 subunități, fiecare dintre ele având o greutate moleculară apropiată de cea a IgG. IgM aparține majorității anticorpilor normali - izohemaglutininele, care sunt prezente în serul de sânge în conformitate cu apartenența oamenilor la anumite grupe de sânge. Aceste variante alotipice de IgM joacă un rol important în transfuzia de sânge. Nu traversează placenta și au cea mai mare aviditate. Atunci când interacționează cu antigenele in vitro, acestea provoacă aglutinarea, precipitarea sau fixarea complementului acestora. În acest din urmă caz, activarea sistemului complement duce la liza antigenelor corpusculare.
(IgA) se găsesc în serul sanguin și în secretele de la suprafața mucoaselor. Serul contine monomeri IgA cu o constanta de sedimentare de 7S la o concentratie de 2,5 g/l. Acest nivel este atins până la vârsta de 10 ani. IgA seric este sintetizată în celulele plasmatice ale splinei, ganglionilor limfatici și membranelor mucoase. Nu aglutinează sau precipită antigenele, nu sunt capabile să activeze complementul în mod clasic și, prin urmare, nu lizează antigenele.
Imunoglobuline secretoare din clasa IgA (SlgA). diferă de ser prin prezența unei componente secretoare; este sintetizat de către celule. epiteliul secretor și poate funcționa ca receptor al acestora, și se alătură IgA atunci când acestea din urmă trece prin celulele epiteliale.IgA secretoare joacă un rol semnificativ în imunitatea locală, deoarece împiedică aderarea microorganismelor la celulele epiteliale ale membranelor mucoase ale gurii, intestinelor. , tractului respirator și urinar.
IgD Neclarificat. Se găsesc pe suprafața limfocitelor B și în ser.
IgE Hipersensibilitatea de tip imediat este realizată prin secretarea de mediatori de către mastocite și bazofile după atașarea antigenului. Principala apărare împotriva invaziei helmintice este prin eliberarea de enzime din eozinofile. Nu fixează complementul.
IgG Principalii anticorpi din răspunsul imun secundar. Ele opsonizează bacteriile și promovează fagocitoza. Fixează complementul, favorizând liza bacteriană. Neutralizează toxinele bacteriene și virușii. Treceți prin placentă.
IgA IgA secretorie previne aderența bacteriilor și virușilor la membranele mucoase. Nu fixează complementul.
IgM Sunt primii care sunt sintetizați atunci când intră un antigen. Fix complement.Nu trece prin placenta. Receptorii antigenici de pe suprafața limfocistului B.
Mecanisme genetice pentru formarea specificității imunoglobulinelor și trecerea celulelor la sinteza imunoglobulinelor dintr-o anumită clasă
Structura moleculelor de Ig este caracterizată de o codificare genetică unică. Folosind metodele geneticii moleculare, s-a dovedit că structura moleculei Ig este controlată de un număr mare de gene care au o organizare fragmentată, formează trei grupuri, sunt localizate pe trei cromozomi diferiți și sunt moștenite independent. Primul grup de gene codifică structura primară a lanțului ușor de tip λ, al doilea - lanțul ușor de tip κ, iar al treilea - toate tipurile de lanțuri grele (α, δ, ε, γ și μ). Genele aparținând fiecărui grup sunt situate pe cromozomul corespunzător în imediata apropiere una de cealaltă, dispuse secvenţial și separate prin introni. Regiunea ADN care codifică structura lanțului ușor de tip λ conține 2 segmente V (controlând structura domeniilor V) și 4 segmente C (controlând structura domeniilor C). Între segmentele C și V există un segment J (din engleză join - connecting). Lanțul ușor de tip κ este codificat de câteva sute de segmente V ADN, 4 segmente J și un segment C. Grupul de gene care controlează structura lanțurilor grele este și mai complex. Alături de segmentele V, C și J ale ADN-ului, acestea includ 20 de segmente D (din engleză divercity - diversity). În plus, există un segment M care codifică biosinteza regiunii asociate membranei a moleculei receptorului Ig. Maturarea pre-limfocitelor B este însoțită de rearanjamente în aparatul lor genetic. Are loc convergența arbitrară a fragmentelor individuale de ADN și asamblarea în cromozomii corespunzători ai genelor unice funcționale. Acest proces se numește splicing (din engleză splicing - merging, docking). Segmentele ADN lipsă sunt excluse de la citirea ulterioară. Pro-ARNm este ulterior transcris din gene funcționale și apoi ARNm final care codifică secvența primară de aminoacizi a lanțurilor L și H ale moleculei Ig. În paralel cu splicing, pot apărea mutații punctuale și completarea fără șablon a oligonucleotidelor în anumite regiuni ale segmentelor V ale genelor imunoglobulinei. Aceste regiuni ale ADN-ului sunt numite regiuni hipermutabile. Splicing-ul și mutația genelor Ig sunt aleatorii. Ele apar în fiecare limfocit independent unul de celălalt și sunt unice, ceea ce crește diversitatea domeniilor V de un număr infinit de ori și, în cele din urmă, structurile paratopilor și determinanții antigenici idiotipici ai moleculei Ig. Prin urmare, limfocitele B specifice aproape oricărui antigen există întotdeauna în organism sau pot apărea în orice moment. Această teză formează baza teoriei genetice moleculare a originii diversității specificităților anticorpilor. În timpul răspunsului imun primar, reproducerea limfocitelor B este, de asemenea, însoțită de rearanjamente de recombinare în cadrul genelor imunoglobulinei, dar deja în cadrul segmentelor C. Acest lucru se manifestă printr-o schimbare secvențială a clasei Ig: în stadiile incipiente de diferențiere, limfocitele B sintetizează Ig din clasele M și D, în stadiile ulterioare - clasele G, A sau E (rar).
Paratop și epitop. Natura interacțiunii antigen-anticorp. Afinitate și aviditate
Epitop sau determinant antigenic - o parte a unei macromolecule de antigen care este recunoscută de sistemul imunitar (anticorpi, limfocite B, limfocite T). Se numește partea unui anticorp care recunoaște un epitop paratop. Deși epitopii se referă de obicei la molecule străine unui organism dat (proteine, glicoproteine, polizaharide etc.), regiunile de molecule de sine recunoscute de sistemul imunitar sunt numite și epitopi.
Antigen - reacție anticorp - o interacțiune specifică a anticorpilor cu antigenele corespunzătoare, în urma căreia se formează complexe antigen-anticorp (complexe imune). Adesea, rezultatul final al acestei reacții este legarea toxinelor, imobilizarea bacteriilor virulente și neutralizarea virusurilor.
Reacția a\r-a\t are loc în două faze, care diferă ca mecanism și viteză. 1. legătura specifică a centrului activ al anticorpului cu grupările corespunzătoare ale antigenului sau haptenei. 2. faza nespecifica - reactie observata vizual.
Asocierea unui antigen cu un anticorp este reversibilă; forța de legătură, numită afinitate, poate fi cuantificat prin determinarea constantei de asociere. Există și un termen aviditate anticorpi, care este utilizat pentru a descrie puterea totală a interacțiunii unui anticorp polivalent cu un antigen polideterminant.
Aviditatea IgM și IgG este foarte importantă în diagnostic și permite analiza retrospectivă a bolilor virale. De exemplu, aviditatea mare a IgM primare indică faza acută a bolii și infecția recentă - de la o lună la o lună și jumătate. Urme de concentrații de IgM pot persista în organism, în unele cazuri, până la doi ani.
30
. Obținerea serurilor pentru reacții imunologiceînvitro.
Anticorpi monoclonali
Seruri antitoxice obtinut prin imunizarea repetata (hiperimunizarea) a cailor, din care se poate obtine o cantitate suficient de mare de sange. Imunizarea se efectuează mai întâi cu toxoid, apoi cu toxină. Serul de sânge este purificat din proteinele de balast prin fermentare și dializă
Seruri antibacteriene obţinute prin hiperimunizarea cailor cu vaccinuri adecvate. Utilizarea serurilor antibacteriene este limitată datorită eficienței lor scăzute.
*material de obtinut heterologă imunoglobulinele sunt ser sau plasmă de animale hiperimunizate.
* material de gatit omolog imunoglobulinele servesc ca plasma sanguina umana.
Seruri aglutinante, obținut prin imunizarea animalelor cu microbi, poate conține anticorpi împotriva microbilor înrudiți, adică sunt polivalenți. Pentru a crește specificitatea serurilor, anticorpii de grup sunt îndepărtați din acestea prin metoda de adsorbție Castellani, folosind antigene de grup. Serurile rezultate se numesc adsorbit. Lăsând anticorpi la un singur antigen, se obțin seruri monoreceptoare.
Anticorpi monoclonali- a/t produs de celulele imune aparținând aceleiași clone celulare, adică descende din același precursor de plasmocite. MA pot fi crescute împotriva aproape oricărui antigen natural (în principal proteine și polizaharide) pe care anticorpul se va lega în mod specific. Ele pot fi utilizate în continuare pentru detectarea (detecția) acestei substanțe sau purificarea acesteia. MA sunt utilizate pe scară largă în biochimie, biologie moleculară și medicină. folosit pentru tratarea melanomului, cancerului de sân.
Aglutinare și precipitare. Reacții de aglutinare și precipitare utilizate în biologie și medicină
Reacții de aglutinare
La aceste reacții iau parte antigenele sub formă de particule (celule microbiene, eritrocite și alți antigene corpusculare), care se lipesc împreună cu anticorpii și precipită. Sunt necesare trei componente pentru a stabili o reacție de aglutinare (RA): 1) antigen (aglutinogen) ; 2) anticorp (aglutinină) și 3) electrolit (soluție izotonică de clorură de sodiu). rezultat pozitiv
- prezența fulgilor aglutina,
negativ
- fara fulgi de aglutinat
Reacție de aglutinare extinsă (RA). Pentru a determina AT în serul sanguin al pacientului, pune reacție de aglutinare extinsă (RA). Pentru a face acest lucru, la o serie de diluții de ser sanguin se adaugă un diagnostic - o suspensie de microorganisme ucise sau particule cu Ag adsorbit. Diluția maximă care dă aglutinare Ag, numit titrul serului sanguin.
Reacția aproximativă de aglutinare (RA) Pentru a identifica microorganismele izolate, un RA aproximativ este plasat pe lame de sticlă. Pentru a face acest lucru, o cultură a agentului patogen este adăugată la o picătură de antiser standard de diagnostic (la o diluție de 1:10, 1:20). Dacă rezultatul este pozitiv, se efectuează o reacție detaliată cu diluții crescânde ale antiserului. reacţie considerat pozitiv dacă se observă aglutinare în diluţii apropiate de titrul serului diagnostic.
Reacții directe de hemaglutinare. Cea mai simplă dintre aceste reacții este aglutinarea eritrocitelor sau hemaglutinarea, folosită pentru determinarea grupelor de sânge în sistemul AB0. Pentru a determina aglutinarea (sau absența acesteia), se folosesc antiseruri standard cu aglutinine anti-A și anti-B. Reacția se numește directă, deoarece antigenele studiate sunt componente naturale ale eritrocitelor.
reacție de precipitare- este formarea și precipitarea unui complex de antigen molecular solubil cu anticorpi sub formă de nor, numit precipitat. Se formează prin amestecarea antigenelor și anticorpilor în cantități echivalente. Reacția de precipitare se pune în eprubete (reacție de precipitare inelară), în geluri, medii nutritive etc.
Reacția de precipitare vă permite să determinați prezența unui antigen necunoscut în materialul de testat prin adăugarea unui anticorp cunoscut sau folosind un antigen cunoscut - un anticorp necunoscut. Precipitația se înregistrează mai bine dacă antigenul este suprapus în eprubetă pe anticorp. În acest caz, se observă apariția unui precipitat sub formă de inel - precipitare inelară. Precipitarea inelului se efectuează în tuburi speciale Precipitarea în agar vă permite să determinați toxigenitatea culturilor de difterie.
În cercetarea criminalistică, precipitarea servește la stabilirea speciilor de sânge, organe și țesuturi folosind seruri precipitante specifice.
Imunoelectroforeza, soiurile sale principale
Imunoelectroforeza (IEF)- o metodă de cercetare a compoziției antigenice a materialelor biologice, combinând electroforeza și imunodifuzia. Descrisă pentru prima dată de Grabar și Williams în 1953, metoda a fost modificată în 1965.
O probă de material antigenic este separată prin electroforeză într-un gel (agaroză), în rezultatul căruia se formează zonele caracteristice. În plus, antiserul precipitant este introdus paralel cu zonele de electroforeză, antigenele și antiserul difuzează unul spre celălalt, iar liniile de precipitare apar la punctul de întâlnire al antiserului cu antigenul, formând un arc. După imunodifuziunea și eluarea moleculelor neprecipitante din gel, gelul este colorat cu coloranți speciali (amido black 10B, azocarmină B și alți coloranți, colorarea proteinelor în cazul antigenelor proteice sau sudan black B în cazul antigenelor lipoproteice) . Există și o serie de modificări ale metodei IEF (folosind un antigen pur, folosind un antiser monospecial, metoda Osserman, metoda Geremans IEF. Cu ajutorul acestei metode în imunologia clinică, concentrația de Ig și identificarea mielomului proteinele sunt determinate.
Contra imunoelectroforeza poate fi folosit pentru a determina antigenele care migrează în agar către electrodul încărcat pozitiv. Este utilizat pentru a identifica antigenele virusului hepatitei B și anticorpii corespunzători, anticorpii la ADN în lupusul eritematos sistemic, autoanticorpii la antigenele nucleare solubile în colagenoză și anticorpii (precipitine) la Aspergillus în aspergiloza bronhopulmonară alergică.
electroforeza rachetei- aceasta este o metodă cantitativă care implică introducerea unui antigen într-un gel care conține anticorpi. Linia de precipitare are forma unei rachete, a cărei lungime este determinată de concentrația antigenului. La fel ca contra electroforeza, aceasta este o metodă rapidă, dar din nou antigenul trebuie să se deplaseze către electrodul încărcat pozitiv. Astfel, electroforeza rachetă este potrivită pentru proteine precum albumina, transferrina și ceruloplasmina, în timp ce concentrația de imunoglobuline este de obicei determinată prin imunodifuzie radială simplă.
Una dintre cele mai de succes opțiuni pentru electroforeza rachetei este imunoelectroforeză bidimensională sau transversală Laurela. În același timp, în prima etapă, amestecul de antigene este separat electroforetic într-un gel de agaroză, apoi proteinele separate sunt din nou forțate să difuzeze în gel sub influența unui câmp electric într-un alt
Tipuri de imunoelectroforeză A - imunoelectroforeză simplă; B- contra imunoelectroforeza; B - imunoelectroforeza rachetă; D - imunoelectroforeză bidimensională.
Metode de imunofluorescență
Imunofluorescență constă în folosirea anticorpilor marcaţi cu fluorocrom, mai exact, fracţia de imunoglobulină a anticorpilor IgG. Un anticorp marcat cu un fluorocrom formează un complex antigen-anticorp cu antigenul, care devine disponibil pentru observare la microscop în raze UV care excită luminescența fluorocromului. Reacția de imunofluorescență directă este utilizată pentru a studia antigenele celulare, pentru a detecta virusul în celulele infectate și pentru a detecta bacteriile și rickettsia în frotiuri. Așadar, pentru diagnosticul rabiei, amprentele bucăților de creier ale animalelor suspectate că poartă un virus sunt tratate cu ser antirabic luminescent. Cu un rezultat pozitiv, aglomerări de culoare verde strălucitoare sunt detectate în citoplasma celulelor nervoase. Diagnosticul rapid al infecției cu gripă, paragripa și adenovirus se bazează pe detectarea antigenelor virale în celulele amprentelor din mucoasa nazală.
Metoda imunofluorescenței indirecte este mai utilizată. bazat pe detectarea unui complex antigen-anticorp folosind un ser de anticorpi luminescent anti-lgG și este utilizat pentru a detecta nu numai antigenele, ci și titrarea anticorpilor. Metoda și-a găsit aplicație în serodiagnosticul herpesului, citomegaliei, febrei Lassa. Preparatele cu ser sanguin de testare stratificat sunt plasate într-un termostat la t° 37° pentru formarea complexelor imune și apoi după spălarea reactivilor nelegați, acești complexe sunt detectați cu ser luminiscent marcat împotriva globulinelor umane. Folosind seruri imune marcate împotriva anticorpilor IgM sau IgG, este posibilă diferențierea tipului de anticorpi și detectarea unui răspuns imun precoce prin prezența anticorpilor IgM.
Radioimunotest
Metoda radioimunologică bazat pe utilizarea unui marcaj radioizotop al antigenelor sau anticorpilor. Este cea mai sensibilă metodă de determinare a antigenelor și anticorpilor, folosită pentru determinarea hormonilor, medicamentelor și antibioticelor, pentru a diagnostica boli bacteriene, virale, rickettsiale, protozoare, pentru a studia proteinele din sânge, antigenele tisulare. A fost dezvoltat inițial ca o metodă specifică pentru măsurarea nivelului de hormoni circulanți în sânge. Sistemul de testare a fost un hormon (antigen) marcat cu un radionuclid și un antiser la acesta. Dacă la un astfel de antiser se adaugă un material care conține hormonul dorit, atunci se va lega o parte din anticorpi, odată cu introducerea ulterioară a hormonului titrat marcat, o cantitate redusă din acesta în comparație cu controlul se va lega de anticorpi. Rezultatul este evaluat prin compararea curbelor etichetei radioactive legate și nelegate. Acest tip de metodă se numește reacție competitivă. Există și alte modificări ale metodei radioimunologice.
Radioimunotest. Principiul radioimunotestului (RIA) se bazează pe detectarea unui complex antigen-anticorp în care unul dintre imunoreactivi a fost marcat cu un izotop radioactiv. Izotopii de iod (I-125 și I-131) sunt utilizați în mod obișnuit. Contabilitatea reacției se realizează în radioactivitate descendentă sau crescătoare (în funcție de metoda RIA) folosind contoare speciale de radiații ionizante. Metoda este foarte sensibilă, dar este înlocuită treptat de imunotestul enzimatic, având în vedere nesiguranța lucrului cu izotopi radioactivi și nevoia de echipamente de înregistrare sofisticate.
O variație a imunoelectroforezei este radioimunoforeza.În acest caz, după separarea electroforetică a antigenelor, mai întâi serul imun marcat cu iod radioactiv împotriva antigenilor detectați este turnat într-un canal tăiat paralel cu mișcarea antigenelor din gel, iar apoi imunul. ser împotriva anticorpilor lgG, care precipită complexele formate ale anticorpului cu antigenul. Toți reactivii nelegați sunt spălați și complexul antigen-anticorp este detectat prin autoradiografie.
Test imunosorbent legat
Metode imunoenzimatice sau imunologice enzimatice se bazează pe utilizarea anticorpilor conjugați cu enzime, în principal peroxidază de hrean sau fosfatază alcalină. Pentru a detecta legarea anticorpilor marcați la un antigen, se adaugă un substrat care este degradat de o enzimă atașată la IgG, colorând galben-maro (peroxidază) sau galben-verde (fosfatază). De asemenea, se folosesc enzime care descompun nu numai substratul cromogen, ci și substratul lumogenic. În acest caz, cu o reacție pozitivă, apare o strălucire. La fel ca imunofluorescența, imunotestul enzimatic este utilizat pentru a detecta antigenele din celule sau pentru a titra anticorpii pe celulele care conțin antigen.
Cel mai popular tip de imunotest enzimatic este imunosorbția. Pe un purtător solid, care poate fi celuloză, poliacrilamidă, dextran și diverse materiale plastice, antigenul este adsorbit. Mai des, suprafața puțurilor micropanourilor servește ca suport. Serul sanguin de testat este adăugat în godeuri cu antigenul sorbit, apoi antiserul marcat cu enzima și substratul. Rezultatele pozitive sunt luate în considerare de schimbarea culorii mediului lichid. Pentru a detecta antigenele, anticorpii sunt adsorbiți pe purtător, apoi materialul de testat este introdus în godeuri și este detectată o reacție cu ser antimicrobian marcat cu enzime. Introducerea avidinei și a biotinei în sistemul de reacție contribuie la creșterea sensibilității metodelor imunofluorescente și a imunotestelor enzimatice.
Imunotestul enzimatic (ELISA). Metodele de imunotest enzimatic folosesc imunoreactivi marcați cu enzime. Cel mai utilizat ELISA în fază solidă. Ca fază solidă se folosesc tablete sau mărgele din polistiren sau polivinil, pe care sunt adsorbiți antigeni sau anticorpi. Pentru a detecta anticorpii, un antigen cunoscut este adsorbit în godeurile unei plăci de polistiren. Apoi se introduce serul de testare, in care se doresc sa detecteze anticorpi la acest antigen. După incubare, godeurile sunt spălate pentru a îndepărta proteinele nelegate și li se adaugă anticorpi anti-imunoglobulină marcați cu enzima. După incubare și spălare, substratul specific enzimei și cromogenul sunt adăugate în godeuri pentru a înregistra produsele finale ale degradării substratului. Prezența și cantitatea de anticorpi se apreciază după modificarea culorii și intensității culorii soluției. Metodele ELISA au sensibilitate și specificitate ridicate și sunt cele mai utilizate pe scară largă dintre metodele imunologice pentru diagnosticul clinic și de laborator.Imunoblotting
Imunoblotting folosit pentru a detecta anticorpi la antigenele individuale sau pentru a „recunoaște” antigene din serurile cunoscute. Metoda constă din 3 etape: separarea macromoleculelor biologice (de exemplu, un virus) în proteine individuale folosind electroforeza pe gel de poliacrilamidă; transferarea proteinelor separate din gel pe un suport solid (blot) prin aplicarea unei plăci de gel de poliacrilamidă pe hârtie activată sau nitroceluloză (electroblotting); detectarea proteinelor dorite pe substrat folosind un imunotest enzimatic direct sau indirect. Ca metodă de diagnosticare, imunoblotting este utilizat pentru infecția cu HIV. Valoarea diagnostică este detectarea anticorpilor la una dintre proteinele învelișului exterior al virusului.
Imunoblotting
După separarea unui amestec complex de proteine prin electroforeză pe gel de poliacrilamidă sau agaroză, acestea pot fi transferate din gel pe o membrană de nitroceluloză microporoasă. În plus, antigenele nespecifice legate de membrană pot fi identificate folosind anticorpi marcați. Această metodă a devenit larg răspândită. De exemplu, este utilizat pentru a identifica componente ale neurofilamentelor care sunt separate anterior pe un gel de poliacrilamidă în prezența dodecil sulfatului de sodiu (SDS). Desigur, dacă antigenul este denaturat ireversibil de SDS, atunci această tehnică nu poate fi utilizată. Dacă proteinele antiserice sunt separate prin focalizare izoelectrică și apoi transferate (aceasta se numește blotting) pe o membrană, atunci așa-numitul spectrotip antiser poate fi determinat și folosind antigenul marcat, adică. determinați izotipul anticorpilor care interacționează cu acest antigen.
răspunsuri care implică complimente.
Sistemul de complement- un complex de proteine complexe care sunt prezente constant în sânge. Acesta este un sistem în cascadă de enzime proteolitice conceput pentru protecția umorală a organismului de acțiunea agenților străini, este implicat în implementarea răspunsului imun al organismului.
Completa- un sistem proteic care include aproximativ 20 de componente care interacționează: C1 (un complex de trei proteine), C2, C3, ..., C9, factorul B, factorul D și o serie de proteine reglatoare. Toate aceste componente sunt proteine solubile care circulă în sânge și în lichidul tisular. Proteinele complementare sunt sintetizate în principal în ficat. Cele mai multe dintre ele sunt inactive până când sunt activate fie printr-un răspuns imun (care implică anticorpi), fie direct de către un microorganism invadator.
Reacții care implică complement bazată pe activarea complementului ca urmare a atașării acestuia la complexul antigen-anticorp. Dacă nu se formează complexul antigen-anticorp, atunci complementul se alătură complexului eritrocit-anticorp antieritrocitar, provocând astfel hemoliza (distrugerea) eritrocitelor (reacție de hemoliză radială). Este folosit pentru a diagnostica boli infecțioase, în special sifilis.
RSK se referă la reacții serologice complexe în care, pe lângă antigen, anticorp și complement, este implicat și sistemul hemolitic, care dezvăluie rezultatele reacției.
RSC se desfășoară în două etape:
primul- interacțiunea antigenului cu anticorpul cu participarea complementului și
al doilea- Identificarea gradului de legare a complementului folosind sistemul hemolitic. Acest sistem este format din eritrocite de oaie și ser hemolitic. Eritrocitele sunt tratate - sensibilizate prin adăugarea de ser la ele la o temperatură de 37 ° C timp de 30 de minute. Liza eritrocitelor de berbec sensibilizate are loc numai în cazul atașării la sistemul complement hemolitic. În absența ei, eritrocitele nu se modifică. Rezultatele RSC depind de prezența anticorpilor în serul de testat. Dacă serul conține anticorpi omologi cu antigenul utilizat în reacție, atunci complexul antigen-anticorp rezultat se atașează, leagă complementul. Când se adaugă un sistem hemolitic, în acest caz, hemoliza nu va avea loc, deoarece întregul complement este cheltuit pe legătura specifică a complexului antigen-anticorp. Eritrocitele rămân neschimbate, astfel încât absența hemolizei în eprubetă este înregistrată ca RSK pozitiv. În absența anticorpilor corespunzători antigenului din ser, complexul specific antigen-anticorp nu se formează și complementul rămâne liber. Când se adaugă un sistem hemolitic, complementul se atașează de acesta și provoacă hemoliza globulelor roșii. Distrugerea globulelor roșii, hemoliza lor caracterizează o reacție negativă.
Reacția de hemoliză. Sub influența anticorpilor și a complementului, suspensia tulbure de eritrocite se transformă într-un lichid transparent roșu strălucitor - sânge lac datorită eliberării de hemoglobină. Reacția este utilizată pe scară largă în practica serologică de laborator ca un indicator al adsorbției complementului la stabilirea unui test de fixare a complementului de diagnostic (RCT). Reacție locală de hemolizăîn gel (reacția Jerne). Această reacție este una dintre variantele hemolizei. Vă permite să determinați numărul de celule care formează anticorpi în organele limfoide. Prezența celulelor care secretă anticorpi hemolitici - hemolizine, este determinată de plăcile de hemoliză care apar într-un gel de agar ce conține eritrocite atunci când acestora li se adaugă țesutul limfoid studiat și complementul. Formarea plăcii se observă numai în jurul acelor celule care secretă anticorpi la eritrocite sau la antigenul care a fost adsorbit anterior pe acestea.
reacție de bacterioliză. Reacția de bacterioliză constă în faptul că atunci când un anumit ser imunitar este combinat cu bacteriile vii corespunzătoare omoloage acestuia în prezența complementului, microbii sunt lizați. Reacția de bacterioliză poate fi observată atât in vitro (in vitro), cât și în organismul animal (in vivo). Acest utilizarea reactieiîn diagnosticul holerei. La stadializarea reacției de bacterioliză în eprubete, se combină o cultură de vibrio izolată de la un pacient, ser antiholeric imun specific și complement. Rezultatele sunt luate în considerare după o incubare de două ore într-un termostat la 37°C prin însămânțarea materialului prelevat din eprubetă pe agar peptonă-carne.
Reacții de neutralizare, reacție de opsonizare
Neutralizare (din lat. neutru- nici una, nici alta) - interacțiunea acizilor cu bazele, în urma căreia se formează săruri și apă. Reacțiile de neutralizare sunt adesea exoterme. De exemplu, reacția hidroxidului de sodiu și acidului clorhidric:
HCl + NaOH = NaCI + H2O
În formă ionică, ecuația se scrie după cum urmează:
H + + OH - \u003d H2O.
Există însă și reacții de neutralizare endotermă, precum reacția bicarbonatului de sodiu (bicarbonat de sodiu) și acidului acetic. Opsonizarea înseamnă facilitarea fagocitozei microorganismelor și a altor materiale după ce opsoninele au fost atașate acestora. Opsonizarea. Aceasta este o reacție imunologică care modifică proprietățile de suprafață ale microorganismelor patogene în așa fel încât acestea devin mai susceptibile la fagocitoză. Opsoninele specifice sunt anticorpi direcționați împotriva antigenelor de suprafață bacteriene care promovează fagocitoza prin acoperirea celulei bacteriene. Activitatea opsoninelor specifice este îmbunătățită de unele componente ale complementului, deși anticorpii corespunzători înșiși pot prezenta, de asemenea, o activitate opsonizantă redusă. Capacitatea de a se lega de celulele tisulare pare a fi foarte pronunțată în IgE, care la om sunt responsabile pentru diferite reacții de hipersensibilitate; poate că această capacitate este determinată de activitatea fragmentului Fc din moleculă.
Anafilaxie, șoc anafilactic, boala serului. Mecanismul de apariție a hipersensibilității de tip imediat. Alergie și alergeni
Anafilaxia este o reacție de hipersensibilitate sistemică care pune viața în pericol la un alergen ( reactie alergica tip imediat). Manifestări de anafilaxie: sindrom de detresă respiratorie, mâncărime, urticarie, umflarea mucoaselor, tulburări ale tractului gastro-intestinal (greață, vărsături, durere, diaree), colaps vascular. Orice alergen poate provoca o reacție anafilactică, dar cele mai semnificative sunt: antiser, hormoni, extracte de polen, venin de himenoptere (Hymenoptera), alimente, medicamente, în special antibiotice; unelte de diagnostic. Forme clinice ale reacțiilor anafilactice: șoc anafilactic, angioedem, urticarie, eritem generalizat. Simptomele bolii: frisoane, amețeli, frică de moarte, senzație de greutate în piept, tahicardie, scăderea tensiunii arteriale, umflarea feței, mâncărimi ale pielii, erupții cutanate asemănătoare urticariei, edem laringian, bronhospasm, greață, vărsături, dureri abdominale, scaune moale.
Șoc anafilacticsau anafilaxie- p-tion alergică încetinită. tip, o stare de sensibilitate puternic crescută a corpului, care se dezvoltă odată cu introducerea repetată a unui alergen. Cauza principală a șocului anafilactic a fost pătrunderea otravii în corpul uman. Patogenia se bazează pe o reacție imediată de hipersensibilitate. Semnul comun și cel mai semnificativ de șoc este o scădere acută a fluxului sanguin cu o încălcare a circulației periferice, iar apoi circulația centrală sub influența histaminei și a altor mediatori, secretați abundent de celule. Pielea devine rece, umedă și cianotică. În legătură cu o scădere a fluxului de sânge în creier și alte organe, apar anxietate, pierderea conștienței, dificultăți de respirație și urinarea este perturbată. Boala serului este o afectiune care se dezvolta in timpul tratamentului cu ser imun de origine animala. Este un răspuns imun la introducerea de proteine străine serice, care constă în formarea unui număr mare de anticorpi care le leagă de celulele plasmatice ale oamenilor org-ma. Acest district yavl. caz special de hipersensibilitate de tip III. Anticorpii umani leagă proteinele străine, formând complexe imune. În același timp, fagocitoza și liza dependentă de complement a complexelor antigen-anticorp apar lent, permițându-le să aibă un efect dăunător asupra organismului. Alergia este o reacție inadecvată a organismului la diferite substanțe, manifestată prin contact direct cu acestea. Se vorbește despre alergii atunci când sistemul imunitar intră în acțiune și organismul răspunde printr-o reacție violentă și o apărare exagerată la substanțele care sunt destul de inofensive în sine. Adică, o alergie este o sensibilitate crescută, un răspuns alterat al corpului uman la impactul anumitor factori - alergeni.
Hipersensibilitate de tip întârziat și mecanismele dezvoltării acesteia
În prezent, în funcție de mecanismul de dezvoltare, se obișnuiește să se distingă 4 tipuri de reacții alergice (hipersensibilitate). Toate aceste tipuri de reacții alergice, de regulă, apar rar în forma lor pură, mai des coexistă în diferite combinații sau trec de la un tip de reacție la altul. În același timp, tipurile I, II și III sunt cauzate de anticorpi, sunt și îi aparțin reacții de hipersensibilitate de tip imediat (ITH). Reacțiile de tip IV sunt cauzate de celulele T sensibilizate și aparțin reacții de hipersensibilitate de tip întârziat (DTH). Al patrulea tip (IV) de reacții este hipersensibilitatea de tip întârziat sau hipersensibilitatea mediată celular. Reacțiile de tip întârziat se dezvoltă într-un organism sensibilizat la 24-48 de ore după contactul cu alergenul. În reacțiile de tip IV, rolul anticorpilor este îndeplinit de limfocitele T sensibilizate. Ag, contactul cu receptorii specifici Ag de pe celulele T, duce la o creștere a numărului acestei populații de limfocite și la activarea lor cu eliberarea de mediatori ai imunității celulare - citokine inflamatorii. Citokinele provoacă acumularea de macrofage și alte limfocite, le implică în procesul de distrugere a AG, ducând la inflamație. Clinic, acest lucru se manifestă prin dezvoltarea inflamației hiperergice: se formează un infiltrat celular, a cărui bază celulară sunt celulele mononucleare - limfocite și monocite. Reacția de tip celular stă la baza dezvoltării infecțiilor virale și bacteriene (dermatită de contact, tuberculoză, micoze, sifilis, lepră, bruceloză), a unor forme de astm bronșic infecțios-alergic, respingere a transplantului și imunitate antitumorală.