Escape types. Plant organs: shoot
Embryo either from an axillary or adnexal (adventitious) kidney. Thus, the kidney is a rudimentary shoot. When the seed germinates from the germinal bud, the first shoot of the plant is formed - its main shoot, or first order escape.
From the main shoot are formed side shoots, or second order shoots, and when branching is repeated - of the third order, etc.
Adventitious shoots are formed from adnexal buds.
This is how the system of shoots is formed, represented by the main shoot and side shoots of the second and subsequent orders. The shoot system increases the total area of contact of the plant with the air.
Depending on the function performed, shoots are distinguished as vegetative, vegetative-generative and generative. Vegetative (unmodified) shoots, consisting of a stem, leaves and buds, and vegetative-generative (partially modified), additionally consisting of a flower or inflorescence, perform the functions of air nutrition and provide the synthesis of organic and inorganic substances. In generative (completely modified) shoots, photosynthesis most often does not occur, but sporangia are formed there, the task of which is to ensure the reproduction of the plant (a flower also belongs to such shoots).
The shoot that produces flowers is called flowering shoot, or peduncle(sometimes the term "peduncle" is understood in a narrower sense - as the section of the stem on which the flowers are located).
Main escape organs
A vegetative unmodified shoot is a single plant organ, consisting of a stem, leaves and buds, formed from a common array of meristems (the shoot growth cone) and having a single conducting system. The stems and leaves, which are the main structural elements of the shoot, are often considered as its constituent organs, that is, organs of the second order. In addition, the obligatory affiliation of the escape is the kidneys. The main external feature that distinguishes the shoot from the root is the presence of leaves.
Monopodial branching
Monopodial branching is the next stage in the evolution of shoot branching. In plants with a monopodial type of shoot structure, the apical bud is preserved throughout the life of the shoot. The monopodial type of branching is often found among gymnosperms, it is also found in many angiosperms (for example, in many species of palms, as well as plants from the Orchid family - gastrohilus, phalaenopsis and others). Some of them have a single vegetative shoot (for example, Phalaenopsis is pleasant).
monopodial plants- the term most often used in the description of plants of tropical and subtropical flora, as well as in popular science literature on indoor and greenhouse floriculture.
Monopodial plants can vary significantly in appearance. Among them there are rosette, with an elongated shoot, bushy.
Sympodial branching
In plants with a sympodial type of shoot structure, the apical bud, having completed development, dies off or gives rise to generative run away. After flowering, this shoot no longer grows, and a new one begins to develop at its base. The structure of the shoot in plants with a sympodial type of branching is more complicated than in plants with; sympodial branching is an evolutionarily more advanced type of branching. The word "simpoidal" is derived from the Greek. sym ("together" or "many") and pod ("leg").
Sympodial branching is characteristic of many angiosperms: for example, lindens, willows, and many orchids.
In orchids, in addition to the apical ones, some sympodial orchids also form lateral inflorescences, developing from buds located at the base of the shoot (Pafinia comb). The part of the shoot pressed against the substrate is called the rhizome. It is located, as a rule, horizontally and does not have true leaves, only scaly. A reduced, almost indistinguishable rhizome occurs in many Masdevallia, Dendrobiums and Oncidiums; well distinguishable and thickened - in cattleyas and lelias, elongated - in bulbophyllums and cologins, reaching 10 or more centimeters. The vertical part of the shoot is often thickened, forming the so-called tuberidium, or pseudobulb. Pseudobulbs can be of various shapes - from almost spherical to cylindrical, cone-shaped, club-shaped and elongated, resembling reed stalks. Pseudobulbs are storage organs.
sympodial plants- the term most often used in the description of plants of tropical and subtropical flora, as well as in popular science literature on indoor and greenhouse floriculture.
|
Bulbophyllum grandiflorum |
Cirrhopetalum macraei |
Oncidium dasystyle. Curtis "s botanical magazine vol. 127 ser. 3 nr. 57 tab. 7787, 1901 |
Dendrobium senile. Curtis "s botanical magazine vol. 127 ser. 3 nr. 57 tab. 7787, 1901 |
Evolution of branch types
Shoot modifications (metamorphosis)
The shoot is the most variable in appearance organ of the plant. This is due not only to the general multifunctionality of vegetative organs that arose in the process of evolution, but also to the changes that occur in the process of plant ontogenesis, due to adaptation to a variety of environmental conditions, and in cultivated plants - under the influence of man.
The rhizome is formed either initially as an underground organ (kupena, raven eye, lily of the valley, blueberry), or first as an above-ground assimilating shoot, which then sinks into the soil with the help of retracting roots (strawberry, lungwort, cuff). Rhizomes can grow and branch monopodially (cuff, crow's eye) or sympodially (kupena, lungwort). Depending on the length of the internodes and the intensity of growth, there are long and short rhizomes and, accordingly, long-rhizome and short-rhizome plants.
When branching rhizomes, it is formed curtain elevated shoots connected by sections of the rhizome system. If the connecting parts are destroyed, the shoots are isolated, and vegetative reproduction occurs. The totality of new individuals formed vegetatively is called clone. Rhizomes are characteristic mainly of herbaceous perennials, but are also found in shrubs (euonymus) and shrubs (lingonberries, blueberries).
close to roots underground stolons- short-lived thin underground shoots bearing underdeveloped scaly leaves. Stolons serve for vegetative reproduction, settlement and territory capture. Spare nutrients are not deposited in them.
In some plants (potato, earthen pear), by the end of summer, stolons form from the apical buds of stolons. tubers (Fig. 4.24). The tuber has a spherical or oval shape, the stem is strongly thickened, reserve nutrients are deposited in it, the leaves are reduced, and buds form in their axils. The stolons die off and collapse, the tubers overwinter, and the next year they give rise to new above-ground shoots.
Tubers do not always develop on stolons. In some perennial plants, the base of the main shoot grows tuberous and thickens (cyclamen, kohlrabi cabbage) ( rice. 4.24). The functions of the tuber are a supply of nutrients, experiencing an unfavorable period of the year, vegetative renewal and reproduction.
In perennial grasses and dwarf shrubs with a well-developed tap root that persists throughout life, a kind of organ of shoot origin is formed, called caudex. Together with the root, it serves as a place for the deposition of reserve substances and carries many renewal buds, some of which may be dormant. The caudex is usually subterranean and is formed from short shoot bases that sink into the soil. Caudex differs from short rhizomes in the way it dies off. Rhizomes, growing at the top, gradually die off and collapse at the older end; the main root is not preserved. The caudex grows in width, from the lower end it gradually turns into a long-lived thickening root. The death and destruction of the caudex and the root goes from the center to the periphery. A cavity is formed in the center, and then it can be divided longitudinally into separate sections - particles. The process of dividing an individual of a taproot plant with a caudex into parts is called particulation. There are many caudex plants among legumes (lupins, alfalfa), umbrella plants (femur, ferula), and Compositae (dandelion, wormwood).
Bulb- this is usually an underground shoot with a very short flattened stem - bottom and scaly fleshy succulent leaves that store water and soluble nutrients, mainly sugars. Aerial shoots grow from the apical and axillary buds of the bulbs, adventitious roots form on the bottom ( rice. 4.24). Thus, the bulb is a typical organ of vegetative renewal and reproduction. Bulbs are most characteristic of plants from the families of lilies (lilies, tulips), onions (onions) and amaryllis (daffodils, hyacinths).
The structure of the bulb is very diverse. In some cases, bulbs storing scales are only modified leaves that do not have green plates (Lily Saranka); in others, these are underground sheaths of green assimilating leaves, which thicken and remain in the bulb after the plates die (onion). Bulb axis growth can be monopodial (snowdrop) or sympodial (hyacinth). The outer scales of the bulb consume the supply of nutrients, dry out and play a protective role. The number of onion scales varies from one (garlic) to several hundred (lilies).
As an organ of renewal and reserve, the bulb is adapted mainly to climates of the Mediterranean type - with fairly mild, wet winters and very hot, dry summers. It serves not so much for a safe overwintering, but for experiencing a harsh summer drought. The storage of water in the tissues of onion scales occurs due to the formation of mucus, which can retain a large amount of water.
Corm outwardly resembles an onion, but its scaly leaves are not storage; they are dry and membranous, and reserve substances are deposited in the thickened stem part (saffron, gladiolus).

Rice. 4.24. Underground escape metamorphoses: 1, 2, 3, 4 - sequence of development and structure of the potato tuber; 5 - cyclamen tuber; 6 - kohlrabi tuber; 7 - bulbs of a tiger lily; 8 - onion bulb; 9 - lily bulb; 10 - section of a long rhizome of couch grass.
Not only underground, but also above-ground shoots of plants can be modified ( rice. 4.25). Quite common elevated stolons. These are plagiotropic short-lived shoots, the function of which is vegetative reproduction, resettlement and territory capture. If stolons carry green leaves and participate in the process of photosynthesis, they are called lashes(bone, tenacious creeping). In strawberries, stolons are devoid of developed green leaves, their stems are thin and fragile, with very long internodes. Such more highly specialized stolons for the function of vegetative reproduction are called mustache.
Juicy, fleshy, adapted for the accumulation of water can be not only bulbs, but also above-ground shoots, usually in plants living in conditions of lack of moisture. Water storage organs can be leaves or stems, sometimes even buds. Such succulent plants are called succulents. Leaf succulents store water in leaf tissues (aloe, agave, jughead, rhodiola, or golden root). Stem succulents are characteristic of the American cactus family and African euphorbiaceae. The succulent stem performs a water-reserving and assimilating function; leaves are reduced or turned into spines ( rice. 4.25, 1). In most cacti, the stems are columnar or spherical, leaves are not formed on them at all, but the nodes are clearly visible by the location of the axillary shoots - areola having the appearance of warts or elongated outgrowths with spines or tufts of hairs. The transformation of leaves into spines reduces the evaporative surface of the plant and protects it from being eaten by animals. An example of the metamorphosis of a kidney into a succulent organ is head of cabbage serves as a cultivated cabbage.
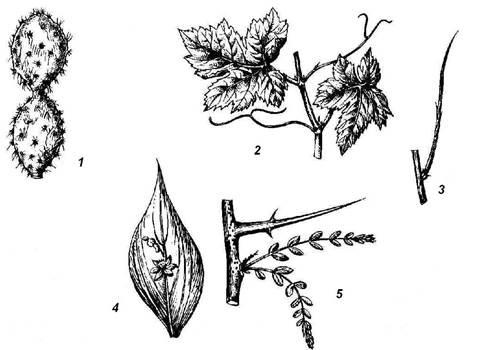
Rice. 4.25. Elevated shoot metamorphoses: 1 - stem succulent (cactus); 2 - tendrils of grapes; 3 - leafless photosynthetic shoot of gorse; 4 - phyllocladium of butcher's broom; 5 - thorn of honey locust.
spines cacti are leafy. Leaf spines are often found in non-succulent plants (barberry) ( rice. 4.26, 1). In many plants, spines are not of leaf, but of stem origin. In the wild apple tree, wild pear, laxative joster, shortened shoots metamorphosed into spines, having limited growth and ending in a point. They acquire the appearance of a hard lignified thorn after the leaves fall. At the hawthorn ( rice. 4.26, 3) the spines that form in the axils of the leaves are completely leafless from the very beginning. In honey locust ( rice. 4.25.5) powerful branched spines are formed on trunks from dormant buds. The formation of spines of any origin, as a rule, is the result of a lack of moisture. When many thorny plants are grown in an artificial humid atmosphere, they lose their spines and instead grow normal leaves (camel thorn) or leafy shoots (English gorse).
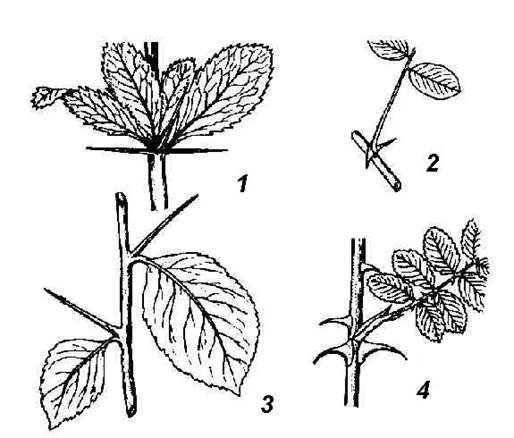
Rice. 4.26. Spines of various origins: 1 - barberry leaf spines; 2 - spines of white acacia, modification of stipules; 3 - spines of hawthorn shoot origin; 4 - thorns - rosehip emergents.
The shoots of a number of plants bear spikes. Thorns differ from spines in smaller sizes, these are outgrowths - emergents - of the integumentary tissue and tissues of the stem bark (rose hips, gooseberries) ( rice. 4.26, 4).
Adaptation to a lack of moisture is very often expressed in the early loss, metamorphosis or reduction of leaves that lose the main function of photosynthesis. This is compensated by the fact that the stem takes on the role of the assimilating organ. Sometimes such an assimilating stem of a leafless shoot remains externally unchanged (Spanish gorse, camel thorn) ( rice. 4.25, 3). The next step in this change of functions is the formation of such organs as phyllocladia and cladodia. These are flattened leaf-like stems or whole shoots. On the shoots of the needle ( rice. 4.25, 4), in the axils of scaly leaves, flat leaf-shaped phylloclades develop, which, like a leaf, have limited growth. Scale-like leaves and inflorescences are formed on phylloclades, which never happens on normal leaves, which means that the phyllocladium corresponds to a whole axillary shoot. Small, needle-like phylloclades are formed in asparagus in the axils of the scaly leaves of the main skeletal shoot. Cladodia are flattened stems that, unlike phyllocladia, retain the ability for long-term growth.
Some plants are characterized by the modification of leaves or their parts, and sometimes entire shoots in antennae, which twist around the support, helping the thin and weak stem to maintain an upright position. In many legumes, the upper part of the pinnate leaf (peas, peas, rank) turns into antennae. In other cases, stipules (sarsaparilla) turn into antennae. Very characteristic tendrils of leafy origin are formed in gourds, and all the transitions from normal to fully metamorphosed leaves can be seen. Antennae of shoot origin can be observed in grapes ( rice. 4.25, 2), passionflower and a number of other plants.
The aerial part of plants is called the term "escape". Its structure is determined by the functions it performs. Of course, each organ is irreplaceable and determines the possibility of the existence of a biological species. Nutritional functions, growth processes, the ability to adapt - these are just some of the most important functions of the visible part of plant organisms.
Biology: the structure of the shoot
In morphology, the axial and lateral parts of this organ are distinguished: the stem and the leaf. The structure is striking in its diversity: from microscopic water duckweed to giant forest sequoia. It is due to the different structure of the constituent parts of the aerial part, associated with the characteristics of the habitat and climatic conditions. There are also shortened rudimentary shoots - buds.
The place where the leaf is attached to the axial part is called the node, and the angle that forms between them is called the sinus. Here are specialized buds that form leaves or flowers. The distance between two leaf attachment points is called an internode.
Stem
The structure of the shoot initially depends on the direction of growth and location in space of the stem. Depending on these features, erect, creeping, creeping, curly and clinging species are distinguished. The stems and the nature of the surface are varied. It can be naked or with outgrowths, smooth or rough. If you cut the stem across, you can determine the shape: rounded, ribbed, with a certain number of faces or flattened.
![]()
Strawberry mustache is also its shoot, which has shortened internodes.
Depending on the life form, grassy and woody stems are distinguished. The first do not have a cambium - lateral In the first years of life, outwardly, new shoots of trees and shrubs look like them. They are green in color and capable of photosynthesis. Over time, they become woody, becoming more durable. They are able to hold large fruits and withstand strong gusts of wind.
stem types
Depending on the characteristics of the development cycle, plants can be one-, two-, and perennial. For example, asters bloom in autumn, after which they completely die off. Carrots and beets develop differently. In the first year of life, they form roots, which are organs that store nutrients. In autumn, their stem dies. But the plant exists in the form of a modified stem. With the onset of favorable conditions, the shoot grows again. At the same time, in the second year of life, as a result of flowering, seeds are formed on it, with the help of which the plant reproduces.

We will consider the life cycle of perennial plants by studying the structural features of the shoots of coniferous plants. These are shrubs or trees that have a single powerful stem - the trunk. Its development begins with the germination of the seed. As a result of its growth, a seedling is formed, and then an adult plant. The life cycle of perennial plants ends with death. Conifers are rightfully considered real centenarians. So, pine lives for about 400 years, spruce - up to 500, and juniper - as many as 1000!
Sheet
The lateral part of the shoot is no less functional and diverse. It provides air nutrition, transpiration - the evaporation of water from the surface, vegetative reproduction. The shoot, the structure of which is determined by the functions performed, is characterized by a variety of leaves.
Cactus needles are needed to reduce the amount of evaporated moisture. And the wide palmate ones of the horse chestnut, on the contrary, increase its quantity.
Leaves with one blade are called simple, and with several, located on the same petiole, they are called complex. Looking at them, you can see a certain pattern. It is created by veins. These are vascular-fibrous bundles. By the nature of venation, leaves are distinguished with mesh (maple, apple), parallel (corn, rye) and arc (plantain, lily of the valley) venation.
leaf arrangement
The shoot, the structure of which also depends on the amount of solar energy received, is characterized by a different arrangement of leaves on the stem. In the case of their arrangement in a spiral, another is formed, and if in a circle - opposite, or whorled.
In nature, there are no plants that do not renew foliage. It is shed by both pine and spruce. Since not all leaves fall at once, no one notices this.

Escape modifications
If it becomes necessary to perform additional functions, the shoots and their components are modified. The leaves may turn into spines or scales. In plant predators, they are able to capture and digest small insects.

The tubers of the Jerusalem artichoke, which is also called the earthen pear, also form a modification of the shoot - a tuber. On a thickened fleshy stem there are scar-like buds from which young shoots grow.
Underground stems with elongated internodes are rhizomes. They look like whips, have well-developed mechanical and conductive tissues. From the buds located on the rhizome, leaves are formed. Those who are new to the structure of rhizomes believe that if you get rid of the leaves, the whole plant will die. But this opinion is erroneous, because the main part of the plant is reliably protected and is underground.
The relationship of structure and functions
The structure of the escape depends on the functions performed. This can be proved by considering the structural elements of its parts. The leaf is covered on the outside with a living tissue skin, in which there are holes - stomata. They are essential for protection and gas exchange. The internal content of the leaf is represented by the main storage and chlorophyll-bearing tissue, which is responsible for the autotrophic nutrition of the entire plant. Conductive and vein-forming elements are the basis for transporting the entire range of essential nutrients.

They are the root (its underground part) and the shoot. The structure of the generative parts does not allow them to perform such functions. They carry out the process of sexual reproduction and distribution of plants. But the flower develops precisely on the stem, and its development requires organic substances formed in the leaves.
It can be concluded that the plant is a single organism, the functioning of the parts of which are interconnected.
Deployment of the shoot from the kidney and its development. There are 2 stages in the life of an escape. The period of formation of the shoot as a rudimentary formation is called intrarenal or embryonic. When the kidney is deployed, the embryonic period in the life of the shoot is replaced by extrarenal or postembryonic period. With the onset of spring, the buds begin to grow and new shoots grow (stems with leaves and buds). You can observe the process of the shoot unfolding even before the onset of spring by placing branches of trees or shrubs in the water (especially in the second half of winter). The deployment of the shoot begins with swelling of the kidney, the kidney scales move apart, the rudiments of green leaves increase in size. Shortly after bud germination, the bud scales fall off, and the remaining scars from fallen scales form a bud ring on the shoot. By long-lasting kidney scars, you can determine the age of a branch of a tree or shrub. At the same time, an elongation of internodes is observed due to actively dividing cells of the interstitial meristem. During this period, intensive growth of leaf blades from the morphologically upper side occurs, and the leaf is folded away from the stem. Due to the intercalated growth, a petiole is formed between the base of the leaf and the blade. The laying of lateral shoots occurs both inside the maternal bud and during the period of extra-bud growth of the shoot.
The concept of annual and elementary shoots.annual shoot - shoot, the growth and formation of which in the extrarenal period of life ends within one year. In a seasonal climate, this occurs in one growing season. The intensity of growth and development of individual metamers is different. Often, at the base of the shoot, the internodes are short and the nodes are close together; higher along the shoot, they become longer, and at the apex, a decrease in the length of the internodes is again observed (the maximum sizes of the internodes, leaves, and buds correspond to the median metameres). Development elementary shoot occurs along a unimodal curve in one growth spurt, or one period of visible growth. Often in an oak, when studying a one-year shoot, it can be noted that it was formed as a result of two periods of growth. Thus, it should be said that the annual shoot consists of two elementary shoots. There are no scaly leaves between two elementary shoots, that is, a bud ring is not formed. In plants of a seasonless climate, annual shoots usually consist of several elementary shoots.
Morphological types of shoots. Escapes vary:
1. Along the length of the internodes.elongated- an escape in which internodes are clearly expressed and the nodes are far from each other. shortened- shoots in which the nodes are close together, and the internodes are practically not expressed or absent (plantain). In the same plant, along with elongated shoots, short ones (apple, birch, hairy sedge) can also develop. Usually shortened shoots are characterized by a small annual increase.
In some plants (pine, club moss), annual shoots are usually more than 10 cm long, but have close internodes. Such escapes are better called long(Fig. 6). The shortened shoots of herbaceous plants are called socket(primrose, dandelion, plantain). Half socket shoots (creeping tenacious, cornflower, meadow cornflower, peach-leaved bellflower) are characterized by contiguous nodes in the basal part of the shoot and elongated ones in its middle part. In the region of the inflorescence, the nodes can be either elongated (spreading bell) or contiguous (crowded bell). In shepherd's purse, wild radish and others, as the flowers open, the nodes in the inflorescence become elongated.
2. By functions. In many plants, specialization of shoots is observed. In woody plants, elongated shoots often vegetative(perform growth and trophic function), and shortened - generative. In elm, bean, wolf's bast, green leaves on generative shoots are completely absent. In herbaceous plants, an inverse correlation is often observed. Shortened shoots are vegetative, and elongated shoots are generative (lily of the valley, plantain).
3. Position of shoots in space. Shoots can be upright (or orthotropic), horizontal (or plagiotropic), ascending (or anisotropic), inclined, twisting around a support, clinging to a support (Fig. 4). The diversity of the position of the shoots of different plants in space allows a greater number of species to grow in a given area.
4. By the time of formation of shoots from the kidneys. We have already considered (see Variety of buds) the peculiarities of the formation of enrichment shoots (sylleptic), renewal shoots and water shoots.
Escape system formation. The formation of the shoot system occurs due to their branching and growth. Branching of a shoot is a process leading to the formation of a shoot on a mother shoot, that is, shoots of the next order are formed on a shoot of one order.
There are two types of shoot branching: 1) apical, 2) lateral. In the process of historical development of plants, the nature of branching changed. Apical, or dichotomous branching is characteristic of club mosses, some ferns, and individual seed plants (some palm trees). Equally dichotomous (equally forked) branching is distinguished - the emerging shoots are the same and unequally dichotomous (unequally forked) - one shoot turns out to be more powerful and is, as it were, a continuation of the maternal shoot.
Rice. 4. Types of shoots by location in space: 1) - orthotropic (Fischer's carnation); 2) - plagiotropic (monetary loosestrife); 3) - anisotropic (moss); 4) - curly (field bindweed); 5) - clinging (mouse peas); 6) - inclined (drooping birch)
The vast majority of seed plants are characterized by lateral branching. Lateral buds give rise to a new shoot. The formation of side shoots increases their total number. The total surface of the air nutrition organs increases, which is extremely important for plants leading an “attached” lifestyle.
Due to the apical bud, the shoot grows in length. Over a number of years, as a result of the activity of one meristem, perennial axes of the same order are formed. This type of growth is called monopodial(Fig. 5). This is how maple, spruce and others grow. However, in a number of plants, the apical meristem forms an inflorescence at a certain stage, and further monopodial growth becomes impossible.
In some species (birch, willow, linden), the apical bud or even part of the shoot dies off. The growth of such plants comes from lateral buds. In autumn, after the death of the apical bud and part of the shoot, one of the lateral buds becomes apical in position, but the presence of a scar from the dead apex of the shoot (branch scar) indicates that this bud is lateral. This type of growth is called sympodial.
Thus, two ways of forming perennial axes are typical for seed plants: 1) monopodial growth and lateral branching (maple, oak, ash), 2) sympodial growth and lateral branching (birch, willow, linden).
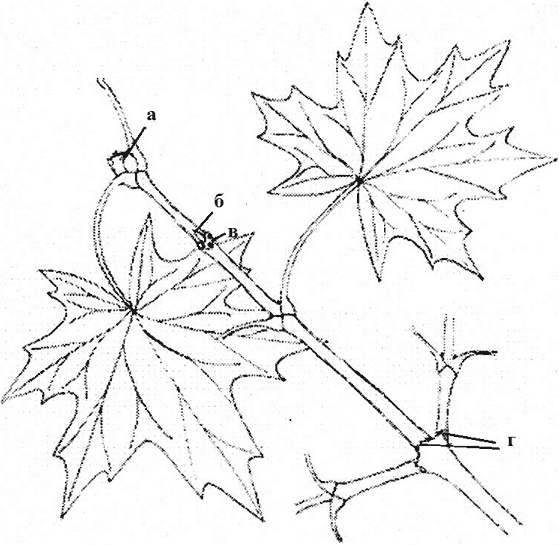
Rice. 5. Monopodial growth of perennial axes with lateral branching. American maple branch (2 years): a - apical kidney; b - axillary kidney; c - leaf scar; d - renal ring
Main and side shoots. The main shoot is formed during the germination of seeds from the embryonic bud. From it in the same year (or in subsequent years) a system of shoots begins to form. From the apical bud there is an increase in the main shoot, the shoot of the first order. From the lateral buds, as a result of lateral branching, lateral shoots are formed, shoots of the second order. Lateral shoots of the second also grow and branch, forming shoots of the third order, and so on.
Acrotonia, mesotonia, basitonia. These three variants of shoot branching are distinguished depending on the location of the most strongly developed side shoots on the mother. At acroton(Greek acros - top, tonos - strength, power) branching, the most powerful side shoots are formed at the top of the mother shoot, with mesoton(Greek mezos - middle) - in the middle, and when basitonic(Greek basis - base) - at its base. A special case of lateral branching is the tillering of the shoot. In this case, side shoots are formed from buds located on a shortened part at the base of the shoot.
Formation of the trunk and crown of trees. Trees are characterized by the formation of a single trunk, usually in its upper part there is intensive branching (acrotonic), which leads to the formation of a crown. Stem growth can be either monopodial or sympodial. In the latter case, the trunk is formed as a result of the activity of the lateral buds by origin. The apical buds, and more often the small upper part of the shoot, are poorly developed and quickly die off. Crown formation occurs due to axillary buds and is associated with different branching intensity. The angle of inclination of the lateral branches relative to the trunk also significantly affects the originality of the crown shape. Usually the first lateral branches are weaker and die off quickly. So in spruce, the formation of full-fledged branches of the crown begins only from 6–8 years, and sometimes even later. Often the shape of the crown directly depends on the growing conditions of the plant. Trees standing alone have a much less developed trunk and a more powerful crown. In a dense forest, trees form a tall trunk and a small crown at the very top.
Shrub formation. Shrubs form several stems that replace each other as they age. The formation of new stems occurs due to dormant buds located at the base of the maternal stem. They can be located both surface and underground. The growth of the trunk occurs over several years. Branching occurs due to axillary buds. The degree of branching is different in different species and often depends on the phytocenosis. If the total life span of a shrub can reach several hundred years, then the stems live for about 20–40 years. However, this value varies widely: from 2 for raspberries to 60 for caragana.
Formation of the shoot system in herbs. Herbaceous plants are characterized by a wide variety of shoot systems, which are formed as a result of lateral branching and monopodial or sympodial growth. Usually, most of the annual growth of grasses dies off in the year of formation. Perennial shoot systems are usually located in the soil or tightly pressed against it. The type of growth and the length of annual growth are of the greatest importance in characterizing the shoot systems of herbaceous plants. Based on these features, shoot formation models perennial herbs (long-shoot sympodial - long-leaved veronica, kupena officinalis; semi-rosette sympodial - spiky veronica, peach-leaved bellflower; rosette monopodial - large plantain, medicinal dandelion; long-shoot monopodial - monetized loosestrife, medicinal veronica).
The concept of monocarpic escape. Monocarpic (mono - one, karpos - fruit) shoot blooms and bears fruit once. The concept of a monocarpic shoot is usually used to characterize herbaceous plants. The fate of a monocarpic shoot in different plants can develop in different ways:
1. The shoot transitioning to flowering in the first year of its development is monocyclic (many-flowered kupena, European hoof).
2. A shoot that starts flowering only in the second year of life is a dicyclic shoot (obscure lungwort, wild strawberry, Kashubian buttercup).
3. If the shoot starts flowering only in the third or subsequent years - a polycyclic shoot (round-leaved wintergreen, sheep fescue).
In addition to the above, there are shoots that never go to flowering. They are called shoots with an incomplete development cycle. The reasons for this may be different: 1) unfavorable conditions; 2) age condition; 3) specialization of shoots in one plant. The last group of plants includes monopodially growing shoots of the large plantain, dandelion officinalis.
leaf arrangement- this is the order of the leaves on the stem (Fig. 6). In some plants, only one leaf departs from the node, as, for example, in birch, oak, linden, buttercup. Such a leaf arrangement is called alternate. If there is more than one leaf on a node, it is whorled, its particular case is opposite, in which there are two leaves within the node, usually located opposite each other (opposite), like in maple, elderberry, viburnum, speedwell. In a number of species (crow's eye, anemone, elodea, juniper), three or more leaves depart from the node. In all cases, leaves extending from two neighboring nodes are never located one above the other, but only at an angle to each other. With this leaf arrangement, minimal shading of one leaf by another is achieved. Often in plants there is an unequal growth of petioles and plates and the placement of leaves in the same plane, while a continuous green screen is formed that perceives the incident rays of the sun. This arrangement of leaves in relation to the light source (often in shading conditions) is called leaf mosaic.
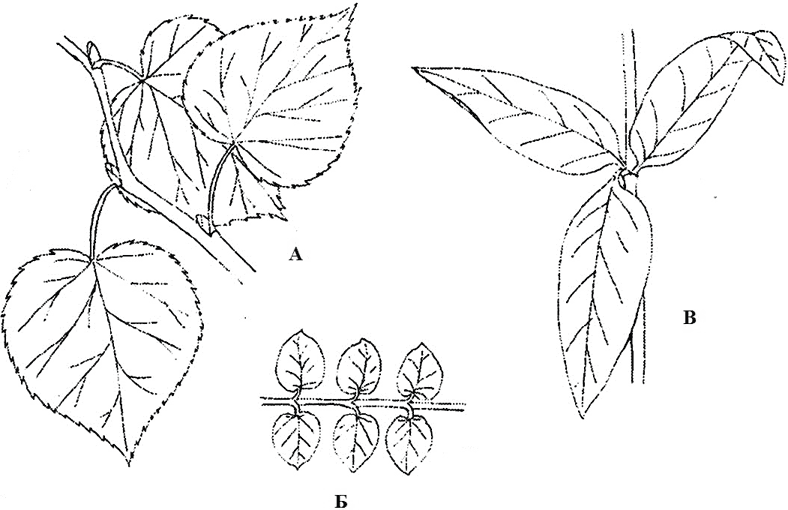
Rice. 6. Types of leaf arrangement: A - next (linden); B - opposite (monetary loosestrife); B - whorled (common loosestrife)
Stem. The central, axial part of the shoot is the stem. The stem performs supporting, transport and storage functions. Green stems are also involved in the air nutrition of plants. The stem is a support for leaves, flowers, fruits, buds and side shoots developing from them. Through the conductive tissues of the stem, from bottom to top and from top to bottom, water and nutrients dissolved in it are transferred. Spare substances are deposited in the tissues of the stem. Young green stems, along with leaves, are involved in the synthesis of organic substances from inorganic ones. Some plants lack green leaves (saxaul, cactus, asparagus, butcher's needle, and others), and the stem is the main organ of air nutrition.
The stem has nodes and internodes. The shape of the stem is usually determined by its cross section, made at the level of the internode. In different plants, it is not the same, but is constant for a species or even a genus, family. This often has taxonomic significance. More often the stem is rounded with a smooth or ribbed edge. It can be tetrahedral (nettle, sage), trihedral (sedge), winged (forest rank), etc. The stem is smooth or pubescent, which is determined by the presence of various hairs on the epidermis.
Sheet is a lateral organ of the shoot, located on the stem. Leaf functions: 1) photosynthesis, 2) transpiration, 3) gas exchange. You will learn more about these concepts in the course of plant physiology.
The main parts of the sheet are plate, petiole, stipules and base(Fig. 7). Their structure corresponds to the functions that the leaf performs, however, in different plants they are not the same in shape and size (Fig. 8). A lamina is an expanded, lamellar part of a leaf. It is this part of the sheet that performs the functions listed above. With a lamellar form of the organ, its maximum surface is achieved, and as a result, high photosynthetic activity. At the base, the plate passes into a stem-like petiole. Its main function is to place the leaf blade in the most favorable position for the plant in space, as well as to ensure the springiness of the leaf, that is, to prevent damage to the leaf during various impacts. It, in turn, in the lower part passes into the base of the leaf, which is directly connected with the stem. The base is an obligatory part of the sheet. In some plants (carrots, wheat), it grows and covers the stem above the node. This basis is called vagina.
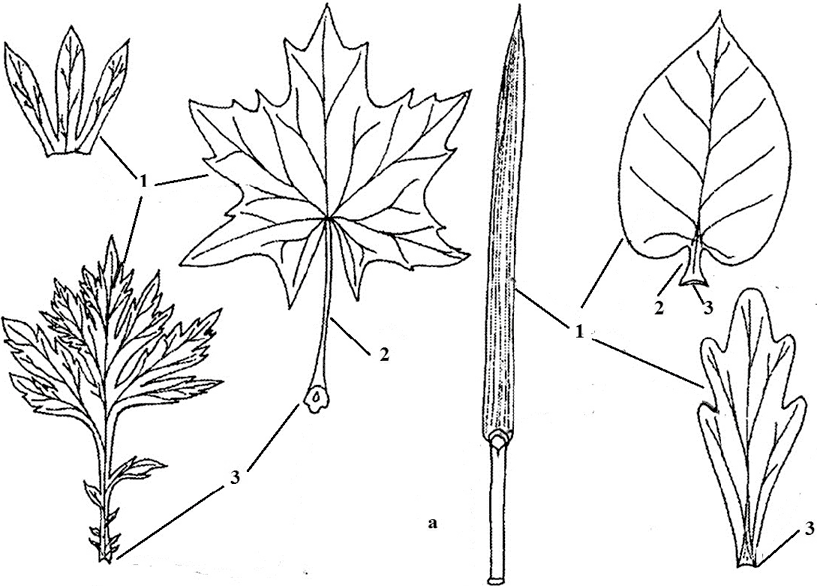
Rice. 7. Simple Leaves: 1 - leaf blade; 2 - petiole; 3 - base; 4 - stipules; a - vagina; b - tongue; c - bell
Stipules are outgrowths at the base of the leaf. Their function is mainly associated with the protection of the leaf blade during the period of intrarenal development. However, in some plants, stipules are able to perform independent functions even in adulthood. They can grow significantly like a pea and resemble a plate, while performing a photosynthetic function. In yellow acacia, gooseberry, stipules turn into spines and serve as protective formations. A leaf is called complete if it has a blade, petiole, base, stipules. A full sheet of mountain ash, rose, oak, bird cherry. In the first two of these plants, all parts of the leaf are preserved throughout life. In oak, adult leaves do not have stipules, as they die early, having performed the functions of protecting the plate of the rudimentary leaf of the kidney. When deploying a kidney and forming a shoot, the stipules of oak, birch, linden and a number of other plants fall off.

Rice. 8. Features of leaf morphology: 1 - parts of the sheet: a - complete sheet; b - compound leaf with stipules; c - stipules fused with petiole; g - bell; d - false leaf arrangement (for example, in bedstraws), f - swollen sheath, g - tubular sheath (for example, in cereals); 2 - position of the leaf on the stem: a - long-leaf, b - short-leaf, c - sessile, d - decurrent, e - stalk-bearing, f - pierced, g - fused leaves; 3 - the shape of the base of the leaf blade: a - wedge-shaped, b - rounded, c - heart-shaped, d - cut, e - swept, f - spear-shaped, g - unequal, h - narrowed; 4 - shape of the leaf apex: a - obtuse, b - truncated, c - sharp, d - pointed, e - pointed, f - notched; 5 - the shape of the edge of the sheet: a - entire, b - serrated, c - double-toothed, d - palmate, e - double-serrate, e - unequally serrated, g - crenate, h - notched, i - wavy, j - ciliated
A leaf is called incomplete if it lacks at least one of its parts: petiole (sessile leaf), stipules or lamina. Sessile leaf of aloe, peach-leaved bell, Fisher's carnation. These plants also lack stipules. The latter are not present in lilac, cabbage, potatoes. Rarely, the plate may be missing. Then its functions are performed by other parts: stipules (leafless rank), flattened petiole (in some acacias).
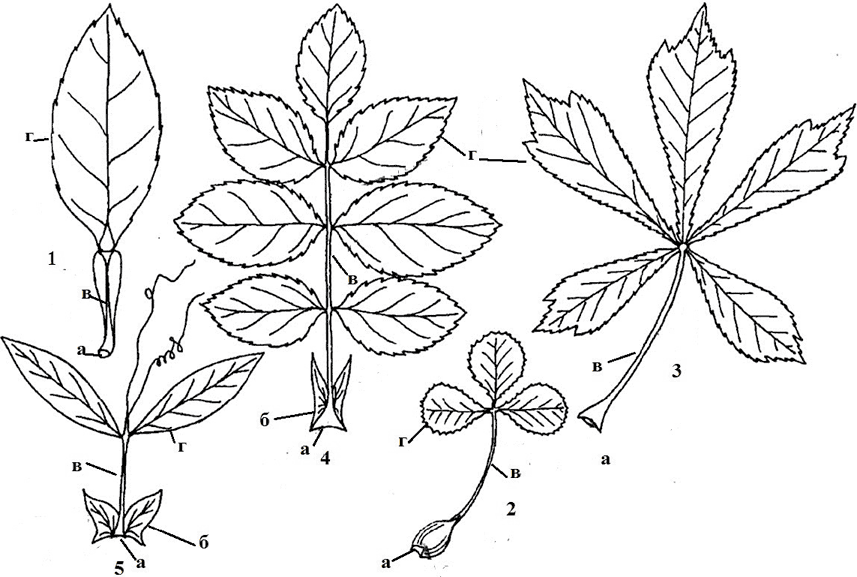
Rice. 9. Compound Leaves: 1 - single leaf (lemon); 2 - ternary (species name); 3 - fingered (horse chestnut); 4 - paired (meadow rank); 5 - unpaired (forest raspberry); a - base; b - stipules; c - rachis; d - leaflet; d - petiole; e - stipules
Simple and compound leaves. A leaf with one blade that does not have an articulation with a petiole or base is called simple. Leaf called difficult(Fig. 9), if it has one or more plates, each of them has its own articulation with a common petiole - the rachis. Each leaf blade of a complex leaf is called a leaflet, or platelet.
Rice. 10. Types of sheet division
A single-leaf compound leaf - in lemon, tangerine, a three-leaf compound leaf - in strawberries, clover, a palmate leaf - in lupine, horse chestnut, an odd-pinnate leaf - in mountain ash, ash (the upper leaflet is one, and only the lateral leaves are arranged in pairs on a common petiole) and paripinnate - in the nomadic, pea (all leaves occupy a lateral position on a common petiole and are arranged in pairs).
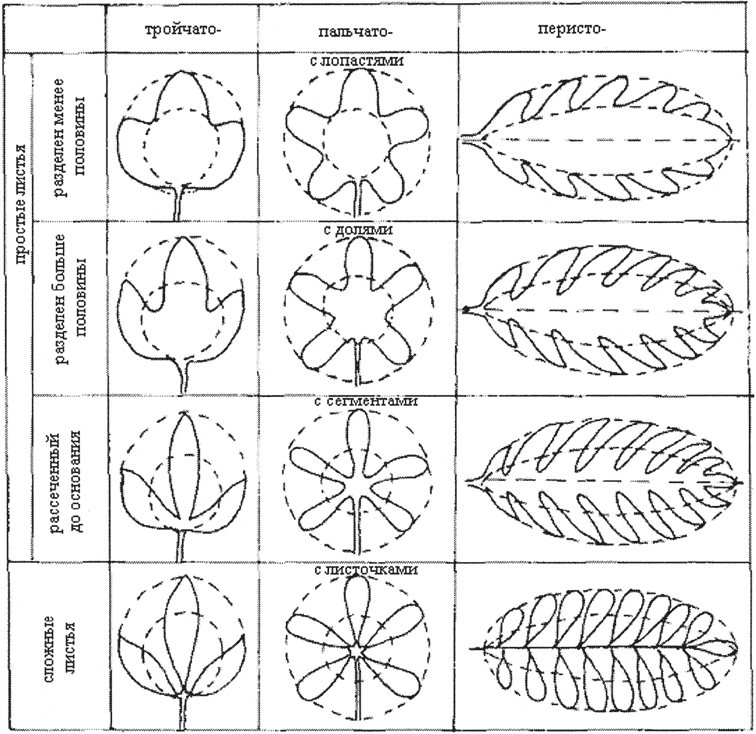
Compound leaves are often confused with simple ones (Fig. 10, 11), which have a deeply dissected plate: triple-dissected - in anemone, palmately dissected - in erect cinquefoil, unpaired pinnate - in goose cinquefoil, lyre-shaped leaf - in potatoes (leaf unpaired pinnate with the largest upper segment) . Each individual part of the plate is called a segment. The segment has no articulation with the petiole. The shape and size of leaves is an important taxonomic feature.
Variety of leaf blades. The blades of simple leaves and leaflets of compound leaves are very diverse in general outline (round, oval, ovate, linear, and others), in the shape of the edge of the blade (the edge can be solid, serrated, serrate, wavy), and in the nature of venation (Fig. 11).
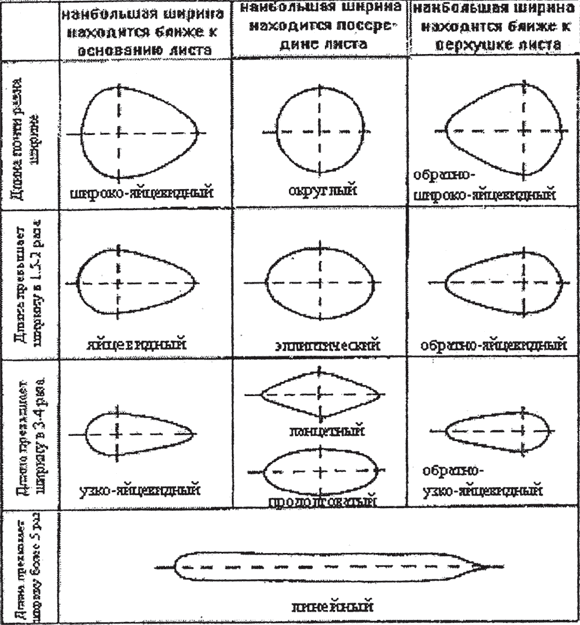
Rice. 11. Blade shapes
Numerous veins cross the plate in different directions. The plate may have one powerful vein running along its middle. it midrib. Thinner lateral branches extend from it to the sides, which in turn branch repeatedly (birch, oak). Such venation of the plate is called pinnate (or pinnate-reticulate). In the presence of several large, more or less identical veins, brought together at the base of the plate and diverging like a fan (geranium, ranunculus), the venation is called palmate (or palmate-reticulate). If large veins run parallel to each other along the plate, then the venation is called parallel (wheat, fescue). Arcuate venation is observed in leaves (lily of the valley, plantain), large veins, in addition to the central one, are curved like an arc (Fig. 12).
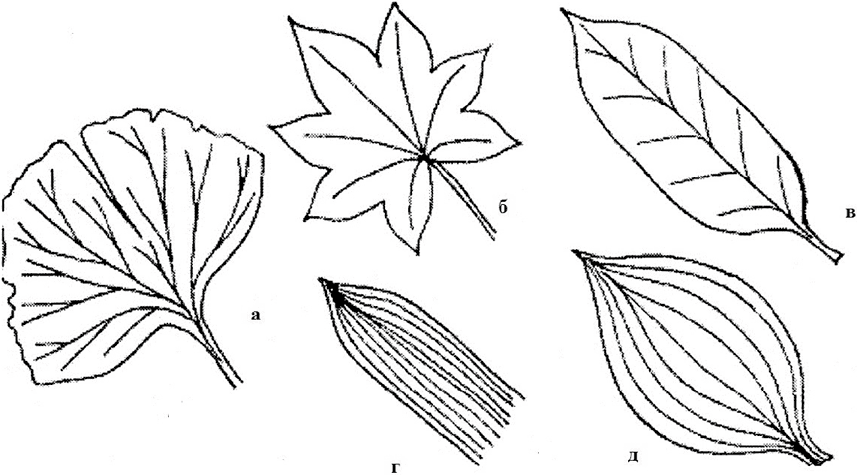
Rice. 12. Forms of leaf venation: a - dichotomous; b - palmate; c - pinnate; d - parallel; d - arc
Three leaf formations. At the base of the annual shoot there are leaves of the lower formation (bud scales, bulb scales), which perform a protective function. Usually they are scaly or membranous, brown, pale green. Ordinary green leaves form a median formation. The leaves of the top formation are located in the region of the inflorescence, are the covering leaves of flowers and perform a protective function (for buds). In some plants (oak maryannik) they are brightly colored and serve to attract insects.
The escape, like the root, is the main organ of the plant. Vegetative shoots typically perform the function of aerial nutrition, but have a number of other functions and are capable of various metamorphoses. spore-bearing shoots (including the flower) are specialized as organs reproductive providing reproduction.
The shoot is formed by the apical meristem as a whole and, therefore, is a single organ of the same rank as the root. However, compared with the root, the shoot has a more complex structure. The vegetative shoot consists of an axial part - stem, which is cylindrical in shape, and leaves- flat lateral organs sitting on the stem. In addition, an obligatory part of the escape are kidneys– rudiments of new shoots, which ensure the growth of the shoot and its branching, i.e. formation of the escape system. The main function of the shoot - photosynthesis - is carried out by leaves; stems are predominantly load-bearing organs that perform mechanical and conductive functions.
The main feature that distinguishes the shoot from the root is its foliage. The part of the stem from which the leaf (leaves) extends is called knot. Stem segments between adjacent nodes internodes. Nodes and internodes are repeated along the axis of the shoot. So the escape has metameric structure, metamer(repeating element) of the shoot are the node with the leaf and the axillary bud and the underlying internode ( rice. 4.16).
Rice. 4.16. Escape structure.
The first shoot of a plant main escape, or escape of the first order. It is formed from an embryonic shoot ending kidney, which forms all subsequent metameres of the main shoot. By position, this kidney is apical; while it persists, this shoot is capable of further growth in length with the formation of new metameres. In addition to the apical, on the shoot are formed lateral kidneys. In seed plants, they are located in the axils of the leaves and are called axillary. From the lateral axillary buds develop lateral shoots, and branching occurs, due to which the total photosynthetic surface of the plant increases. Formed escape system, represented by the main shoot (shoot of the first order) and side shoots (shoots of the second order), and when branching is repeated, by side shoots of the third, fourth and subsequent orders. A shoot of any order has its own apical bud and is capable of growing in length.
Bud- this is a rudimentary, not yet unfolded shoot. Inside the kidney is the meristematic tip of the shoot - its apex(rice. 4.17). The apex is an actively working growth center that ensures the formation of all organs and primary tissues of the shoot. The source of constant self-renewal of the apex is the initial cells of the apical meristem, concentrated at the tip of the apex. The vegetative shoot apex, in contrast to the always smooth root apex, regularly forms protrusions on the surface, which are the beginnings of leaves. Only the very tip of the apex, which is called growth cone escape. Its shape varies greatly in different plants and does not always look like a cone; the apical part of the apex can be low, hemispherical, flat, or even concave.
From vegetative buds develop vegetative shoots consisting of a stem, leaves and buds. Such a kidney consists of a meristematic rudimentary axis ending growth cone, and rudimentary leaves of different ages. Due to uneven growth, the lower leaf primordia are bent inward and cover the upper, younger, leaf primordia and the growth cone. The nodes in the kidney are close together, since the internodes have not yet had time to stretch out. In the axils of leaf rudiments in the kidney, the rudiments of axillary buds of the following order can already be laid ( rice. 4.17). AT vegetative-generative a number of vegetative metameres are laid in the buds, and the growth cone is turned into a rudimentary flower or inflorescence. Generative, or floral the buds contain only the rudiment of an inflorescence or a single flower, in the latter case the bud is called bud.
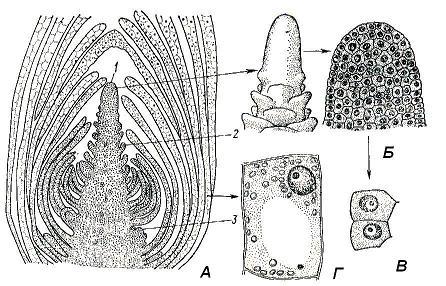
Rice. 4.17. The apical bud of the Elodea shoot: A - longitudinal section; B - growth cone (appearance and longitudinal section); C – cells of the apical meristem; D - parenchymal cell of the formed leaf; 1 - growth cone; 2 - leaf rudiment; 3 - the rudiment of the axillary kidney.
The outer leaves of the bud often change into kidney scales, which perform a protective function and protect the meristematic parts of the kidney from drying out and sudden changes in temperature. Such kidneys are called closed(wintering buds of trees and shrubs and some perennial grasses). open kidneys do not have kidney scales.
In addition to the usual, exogenous in inception, axillary buds, plants often form adnexal, or adventive kidneys. They arise not in the meristematic tip of the shoot, but on the adult, already differentiated part of the organ, endogenously, from internal tissues. Adnexal buds can form on stems (then they are usually located in internodes), leaves and roots. Adnexal buds are of great biological importance: they provide active vegetative renewal and reproduction of those perennial plants that have them. In particular, with the help of adnexal kidneys, they renew and multiply root offspring plants (raspberry, aspen, thistle, dandelion). Root offspring- these are shoots that have developed from adventitious buds on the roots. Adnexal buds on the leaves are formed relatively rarely. If such buds immediately give small shoots with adventitious roots that fall off the mother leaf and grow into new individuals, they are called brood(bryophyllum).
In the seasonal climate of the temperate zone, the deployment of shoots from the buds in most plants is periodic. In trees and shrubs, as well as in many perennial herbaceous plants, buds unfold into shoots once a year - in spring or early summer, after which new wintering buds are formed with the beginnings of next year's shoots. Shoots that grow from buds in one growing season are called annual shoots, or annual increments. In trees, they are well distinguished due to the formation renal rings- scars that remain on the stem after the fall of the kidney scales. In the summer of our deciduous trees, the annual shoots of only the current year are covered with leaves; there are no leaves on the annual shoots of previous years. In evergreen trees, leaves can be preserved on the corresponding annual increments of 3-5 past years. In a seasonally unseasoned climate, several shoots may form in one year, separated by small dormant periods. Such shoots formed in one growth cycle are called elementary shoots.
Buds that fall into a dormant state for a while, and then give new elementary and annual shoots, are called wintering or resting. According to their function, they can be called kidney regular renewal. Such buds are an obligatory feature of any perennial plant, woody or herbaceous, they ensure the perennial existence of an individual. By origin, renewal kidneys can be both exogenous (apical or axillary) and endogenous (adnexal).
If the lateral buds do not have a dormant period and develop simultaneously with the growth of the maternal shoot, they are called kidney enrichment. Deploying ones enrichment shoots greatly increase (enrich) the total photosynthetic surface of the plant, as well as the total number of inflorescences formed and, consequently, seed productivity. Enrichment shoots are typical for most annual grasses and for a number of perennial herbaceous plants with elongated flowering shoots.
A special category is dormant buds, very characteristic of deciduous trees, shrubs, shrubs and a number of perennial grasses. By origin, they, like the buds of regular renewal, can be axillary and adnexal, but, unlike them, do not turn into shoots for many years. The stimulus for the awakening of dormant buds is usually either damage to the main trunk or branch (stump growth after cutting down a number of trees), or natural aging of the maternal shoot system associated with the attenuation of the vital activity of normal renewal buds (change of stems in shrubs). In some plants, leafless flowering shoots form from dormant buds on the trunk. This phenomenon is called caulifloria and is characteristic of many rainforest trees, such as the chocolate tree. In honey locust, bunches of large branched spines grow from sleeping buds on the trunk - modified shoots ( rice. 4.18).
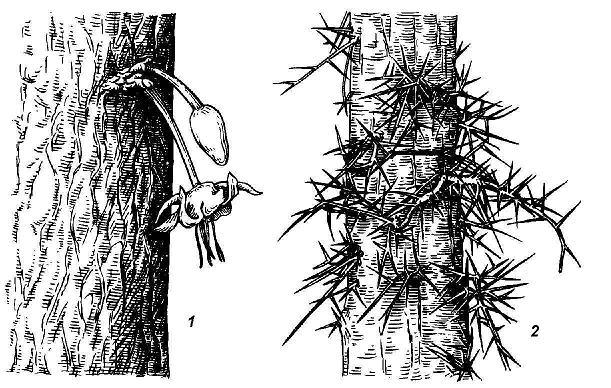
Rice. 4.18. Shoots from dormant buds: 1 - caulifloria near the chocolate tree; 2 - spines in honey locust from branched dormant buds.
Direction of shoot growth. Shoots growing vertically, perpendicular to the surface of the earth, are called orthotropic. Horizontally growing shoots are called plagiotropic. The direction of growth may change during shoot development.
Depending on the position in space, morphological types of shoots are distinguished ( rice. 4.19). The main shoot in most cases retains orthotropic growth and remains upright. Lateral shoots can grow in different directions, often forming a different angle with the parent shoot. In the process of growth, the shoot can change direction from plagiotropic to orthotropic, then it is called rising, or ascending. Shoots with plagiotropic growth that persists throughout life are called creeping. If they form adventitious roots at the nodes, they are called creeping.
Orthotropic growth is connected in a certain way with the degree of development of mechanical tissues. In the absence of well-developed mechanical tissues in elongated shoots, orthotropic growth is impossible. But often plants that do not have a sufficiently developed internal skeleton still grow upward. This is achieved in various ways. Weak shoots of such plants - creeper twist around some kind of solid support ( curly shoots), climb with the help of various kinds of spines, hooks, roots - trailers ( climbing shoots), cling with the help of antennae of various origins ( clinging shoots).
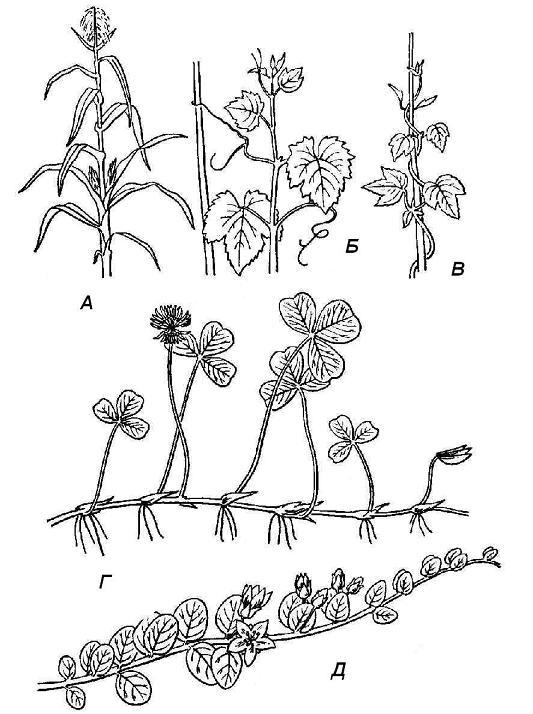
Rice. 4.19. Types of shoots by position in space: A - upright; B - clinging; B - curly; G - creeping; D - creeping.
Leaf arrangement. leaf arrangement, or phyllotaxis- the order of placement of leaves on the axis of the shoot. There are several main types of leaf arrangement ( rice. 4.20).
Spiral, or another leaf arrangement is observed when there is one leaf at each node, and the bases of successive leaves can be connected by a conditional spiral line. double row leaf arrangement can be considered as a special case of spiral. At the same time, at each node there is one sheet, covering the entire or almost the entire circumference of the axis with a wide base. Whorled leaf arrangement occurs when several leaves are laid on one node. Opposite leaf arrangement - a special case of whorled, when two leaves are formed on one node, exactly opposite each other; most often such a leaf arrangement occurs cross opposite, i.e. neighboring pairs of leaves are in mutually perpendicular planes ( rice. 4.20).
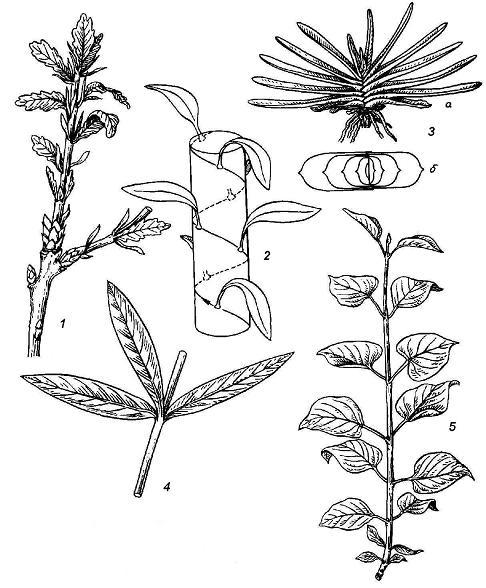
Rice. 4.20. Types of leaf arrangement: 1 - spiral in oak; 2 - scheme of spiral leaf arrangement; 3 - two-row in gasteria ( a- side view of the plant b– top view, scheme); 4 - whorled in oleander; 5 - opposite in lilac.
The order of initiation of leaf primordia on the shoot apex is a hereditary trait of each species, sometimes characteristic of a genus and even an entire family of plants. The leaf arrangement of the adult shoot is determined primarily by genetic factors. However, during the development of the shoot from the bud and its further growth, the location of the leaves can be influenced by external factors, mainly lighting conditions and gravity. Therefore, the final picture of leaf arrangement can differ greatly from the initial one and usually acquires a pronounced adaptive character. The leaves are arranged so that their plates are in the most favorable lighting conditions in each case. This is most pronounced in the form sheet mosaic observed on plagiotropic and rosette shoots of plants. In this case, the plates of all leaves are arranged horizontally, the leaves do not obscure each other, but form a single plane where there are no gaps; smaller leaves fill the gaps between the larger ones.
Shoot branching types. Branching is the formation of a system of axes. It provides an increase in the total area of contact of the plant body with air, water or soil. Branching arose in the process of evolution even before the appearance of organs. In the simplest case, the top of the main axis forks and gives rise to two axes of the next order. it apical, or dichotomous branching. Many multicellular algae have apical branching, as well as some primitive plants, such as club mosses ( rice. 4.21).
Other groups of plants are characterized by a more specialized side branch type. In this case, the lateral branches are laid below the top of the main axis, without affecting its ability to further increase. With this method, the potential for branching and formation of organ systems is much more extensive and biologically beneficial.
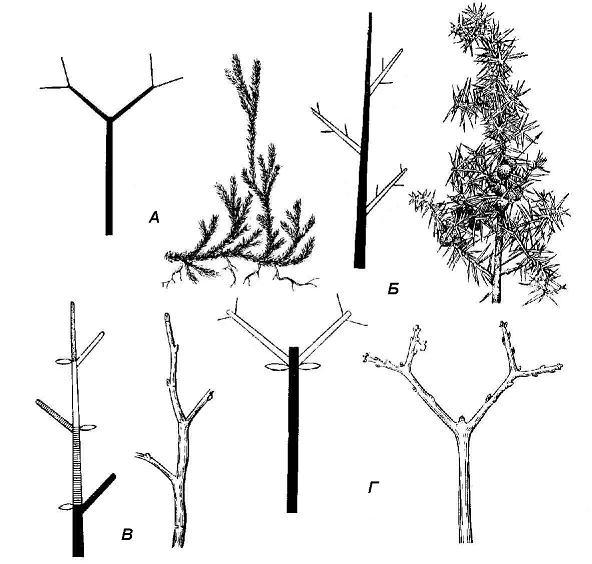
Rice. 4.21. Shoot branching types: A - dichotomous (club moss); B - monopodial (juniper); B - sympodial type of monochasia (bird cherry); D - sympodial according to the type of dichasia (maple).
There are two types of lateral branching: monopodial and sympodial(rice. 4.21). With a monopodial branching system, each axis is a monopodium, i.e. the result of the work of one apical meristem. Monopodial branching is characteristic of most gymnosperms and many herbaceous angiosperms. Most angiosperms, however, branch in a sympodial pattern. With sympodial branching, the apical bud of the shoot dies off at a certain stage or stops active growth, but an increased development of one or more lateral buds begins. Shoots are formed from them, replacing the shoot that has stopped growing. The resulting axis is a sympodium - a composite axis consisting of axes of several successive orders. The ability of plants to sympodial branching is of great biological importance. In case of damage to the apical bud, the growth of the axis will continue with lateral shoots.
Depending on the number of replacement axes, sympodial branching is distinguished by type monochasia, dichasia and pleiochasia. Branching according to the type of dichasia, or false dichotomous branching is typical for shoots with opposite leaf arrangement (lilac, viburnum).
In some groups of plants, the growth of the main skeletal axes occurs due to one or a few apical buds, lateral skeletal branches are not formed at all or are formed in a very small number. Tree-like plants of this type are found mainly in tropical areas (palm trees, dracaena, yucca, agave, cycads). The crown of these plants is formed not by branches, but by large leaves brought together in a rosette at the top of the trunk. The ability to rapidly grow and capture space, as well as to recover from damage in such plants is often absent or weakly expressed. Among temperate trees, such non-branching forms are practically not found.
The other extreme is plants that branch too profusely. They are represented by the life form cushion plants (rice. 4.22). The growth in the length of the shoots of these plants is extremely limited, but on the other hand, many lateral branches are formed annually, diverging in all directions. The surface of the shoot system of the plant looks as if trimmed; some pillows are so dense that they look like stones.
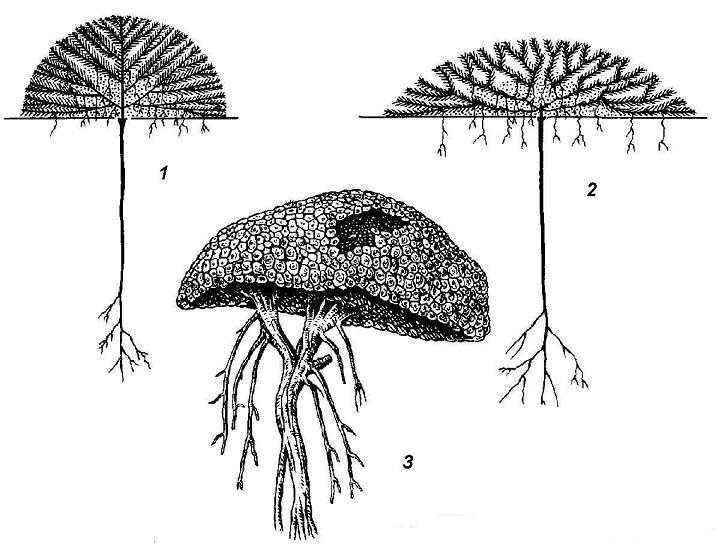
Rice. 4.22. Plants - pillows: 1, 2 - schemes of the structure of pillow plants; 3 - Azorella from Kerguelen Island.
Representatives of a life form branch very strongly Tumbleweed characteristic of steppe plants. A spherically branched, very loose system of shoots is a huge inflorescence, which, after fruit ripening, breaks off at the base of the stem and rolls over the steppe with the wind, scattering the seeds.
Specialization and metamorphoses of shoots. Many plants within the shoot system have a certain specialization. Orthotropic and plagiotropic, elongated and shortened shoots perform different functions.
elongated called shoots with normally developed internodes. In woody plants, they are called growth and are located along the periphery of the crown, determining its shape. Their main function is to capture space, increase the volume of photosynthetic organs. shortened shoots have close nodes and very short internodes ( rice. 4.23). They form inside the crown and absorb scattered light penetrating there. Often shortened shoots of trees are flowering and perform the function of reproduction.
Rice. 4.23. Shortened (A) and elongated (B) sycamore shoots: 1 - internode; 2 - annual increments.
Herbaceous plants usually have shortened rosette shoots perform the function of perennial skeletal and photosynthetic, and elongated ones are formed in the axils of rosette leaves and are flower-bearing (plantain, cuff, violets). If axillary peduncles are leafless, they are called arrows. The fact that flowering shoots are short in woody plants and elongated in herbaceous plants is biologically well explained. For successful pollination, grass inflorescences must be raised above the herbage, and in trees, even shortened shoots in the crown are in favorable conditions for pollination.
An example of the specialization of shoots is the perennial axial organs of woody plants - trunks and branches crowns. In deciduous trees, annual shoots lose their assimilation function after the first growing season, in evergreen trees - after a few years. Some of the shoots die off completely after the loss of leaves, but the majority remain as skeletal axes, performing supporting, conducting, and storage functions for decades. The leafless skeletal axes are known as boughs and trunks(by the trees) stems(for shrubs).
In the course of adaptation to specific environmental conditions or in connection with a sharp change in functions, shoots can change (metamorphize). Shoots developing underground are especially often metamorphosed. Such shoots lose the function of photosynthesis; they are common in perennial plants, where they act as organs for experiencing an unfavorable period of the year, stock and renewal.
The most common underground shoot metamorphosis is rhizome (rice. 4.24). It is customary to call a rhizome a long-lived underground shoot that performs the functions of deposition of reserve nutrients, renewal, and sometimes vegetative propagation. The rhizome is formed in perennial plants, which, as a rule, do not have a main root in the adult state. According to its position in space, it can be horizontal, oblique or vertical. The rhizome usually does not bear green leaves, but, being a shoot, retains a metameric structure. The nodes are distinguished either by leaf scars and the remains of dry leaves, or by living scaly leaves; axillary buds are also located in the nodes. According to these features, the rhizome is easy to distinguish from the root. As a rule, adventitious roots are formed on the rhizome; lateral branches of the rhizome and above-ground shoots grow from the buds.
The rhizome is formed either initially as an underground organ (kupena, raven eye, lily of the valley, blueberry), or first as an above-ground assimilating shoot, which then sinks into the soil with the help of retracting roots (strawberry, lungwort, cuff). Rhizomes can grow and branch monopodially (cuff, crow's eye) or sympodially (kupena, lungwort). Depending on the length of the internodes and the intensity of growth, there are long and short rhizomes and, accordingly, long-rhizome and short-rhizome plants.
When branching rhizomes, it is formed curtain elevated shoots connected by sections of the rhizome system. If the connecting parts are destroyed, the shoots are isolated, and vegetative reproduction occurs. The totality of new individuals formed vegetatively is called clone. Rhizomes are characteristic mainly of herbaceous perennials, but are also found in shrubs (euonymus) and shrubs (lingonberries, blueberries).
close to roots underground stolons- short-lived thin underground shoots bearing underdeveloped scaly leaves. Stolons serve for vegetative reproduction, settlement and territory capture. Spare nutrients are not deposited in them.
In some plants (potato, earthen pear), by the end of summer, stolons form from the apical buds of stolons. tubers (Fig. 4.24). The tuber has a spherical or oval shape, the stem is strongly thickened, reserve nutrients are deposited in it, the leaves are reduced, and buds form in their axils. The stolons die off and collapse, the tubers overwinter, and the next year they give rise to new above-ground shoots.
Tubers do not always develop on stolons. In some perennial plants, the base of the main shoot grows tuberous and thickens (cyclamen, kohlrabi cabbage) ( rice. 4.24). The functions of the tuber are a supply of nutrients, experiencing an unfavorable period of the year, vegetative renewal and reproduction.
In perennial grasses and dwarf shrubs with a well-developed tap root that persists throughout life, a kind of organ of shoot origin is formed, called caudex. Together with the root, it serves as a place for the deposition of reserve substances and carries many renewal buds, some of which may be dormant. The caudex is usually subterranean and is formed from short shoot bases that sink into the soil. Caudex differs from short rhizomes in the way it dies off. Rhizomes, growing at the top, gradually die off and collapse at the older end; the main root is not preserved. The caudex grows in width, from the lower end it gradually turns into a long-lived thickening root. The death and destruction of the caudex and the root goes from the center to the periphery. A cavity is formed in the center, and then it can be divided longitudinally into separate sections - particles. The process of dividing an individual of a taproot plant with a caudex into parts is called particulation. There are many caudex plants among legumes (lupins, alfalfa), umbrella plants (femur, ferula), and Compositae (dandelion, wormwood).
Bulb- this is usually an underground shoot with a very short flattened stem - bottom and scaly fleshy succulent leaves that store water and soluble nutrients, mainly sugars. Aerial shoots grow from the apical and axillary buds of the bulbs, adventitious roots form on the bottom ( rice. 4.24). Thus, the bulb is a typical organ of vegetative renewal and reproduction. Bulbs are most characteristic of plants from the families of lilies (lilies, tulips), onions (onions) and amaryllis (daffodils, hyacinths).
The structure of the bulb is very diverse. In some cases, bulbs storing scales are only modified leaves that do not have green plates (lily saranka); in others, these are underground sheaths of green assimilating leaves, which thicken and remain in the bulb after the plates die (onion). Bulb axis growth can be monopodial (snowdrop) or sympodial (hyacinth). The outer scales of the bulb consume the supply of nutrients, dry out and play a protective role. The number of onion scales varies from one (garlic) to several hundred (lilies).
As an organ of renewal and reserve, the bulb is adapted mainly to climates of the Mediterranean type - with fairly mild, wet winters and very hot, dry summers. It serves not so much for a safe overwintering, but for experiencing a harsh summer drought. The storage of water in the tissues of onion scales occurs due to the formation of mucus, which can retain a large amount of water.
Corm outwardly resembles an onion, but its scaly leaves are not storage; they are dry and membranous, and reserve substances are deposited in the thickened stem part (saffron, gladiolus).

Rice. 4.24. Underground escape metamorphoses: 1, 2, 3, 4 - sequence of development and structure of the potato tuber; 5 - cyclamen tuber; 6 - kohlrabi tuber; 7 - bulbs of a tiger lily; 8 - onion bulb; 9 - lily bulb; 10 - section of a long rhizome of couch grass.
Not only underground, but also above-ground shoots of plants can be modified ( rice. 4.25). Quite common elevated stolons. These are plagiotropic short-lived shoots, the function of which is vegetative reproduction, resettlement and territory capture. If stolons carry green leaves and participate in the process of photosynthesis, they are called lashes(bone, tenacious creeping). In strawberries, stolons are devoid of developed green leaves, their stems are thin and fragile, with very long internodes. Such more highly specialized stolons for the function of vegetative reproduction are called mustache.
Juicy, fleshy, adapted for the accumulation of water can be not only bulbs, but also above-ground shoots, usually in plants living in conditions of lack of moisture. Water storage organs can be leaves or stems, sometimes even buds. Such succulent plants are called succulents. Leaf succulents store water in leaf tissues (aloe, agave, jughead, rhodiola, or golden root). Stem succulents are characteristic of the American cactus family and African euphorbiaceae. The succulent stem performs a water-reserving and assimilating function; leaves are reduced or turned into spines ( rice. 4.25, 1). In most cacti, the stems are columnar or spherical, leaves are not formed on them at all, but the nodes are clearly visible by the location of the axillary shoots - areola having the appearance of warts or elongated outgrowths with spines or tufts of hairs. The transformation of leaves into spines reduces the evaporative surface of the plant and protects it from being eaten by animals. An example of the metamorphosis of a kidney into a succulent organ is head of cabbage serves as a cultivated cabbage.
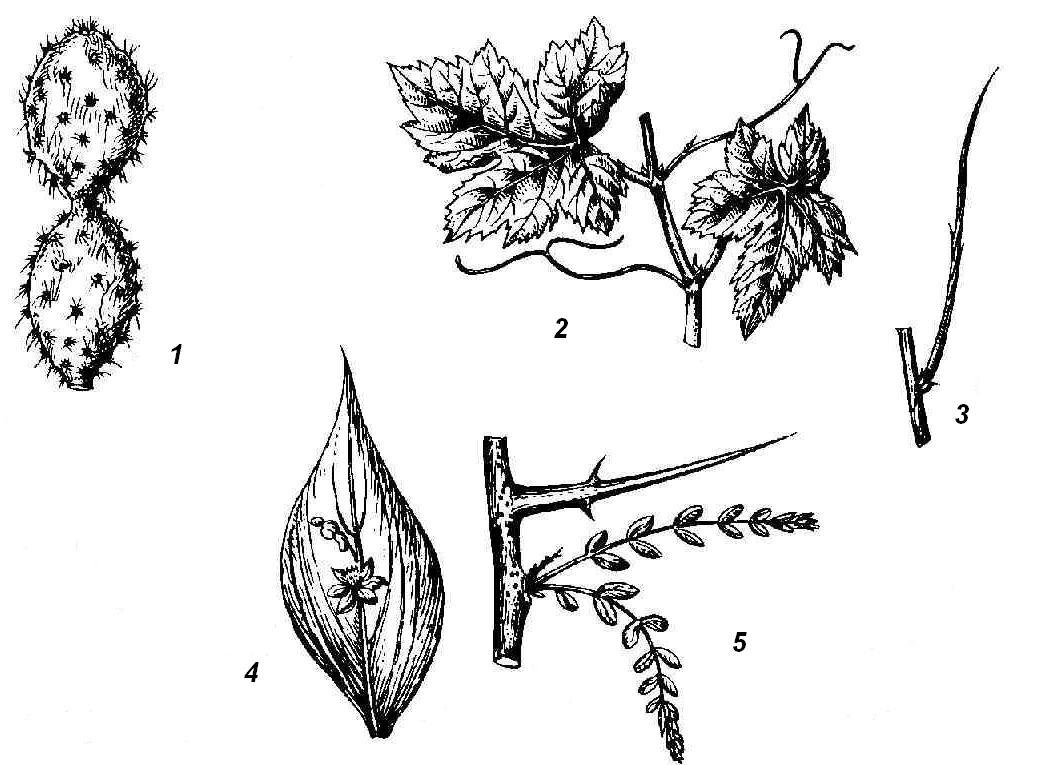
Rice. 4.25. Elevated shoot metamorphoses: 1 - stem succulent (cactus); 2 - tendrils of grapes; 3 - leafless photosynthetic shoot of gorse; 4 - phyllocladium of butcher's broom; 5 - thorn of honey locust.
spines cacti are leafy. Leaf spines are often found in non-succulent plants (barberry) ( rice. 4.26, 1). In many plants, spines are not of leaf, but of stem origin. In the wild apple tree, wild pear, laxative joster, shortened shoots metamorphosed into spines, having limited growth and ending in a point. They acquire the appearance of a hard lignified thorn after the leaves fall. At the hawthorn ( rice. 4.26, 3) the spines that form in the axils of the leaves are completely leafless from the very beginning. In honey locust ( rice. 4.25.5) powerful branched spines are formed on trunks from dormant buds. The formation of spines of any origin, as a rule, is the result of a lack of moisture. When many thorny plants are grown in an artificial humid atmosphere, they lose their spines and instead grow normal leaves (camel thorn) or leafy shoots (English gorse).
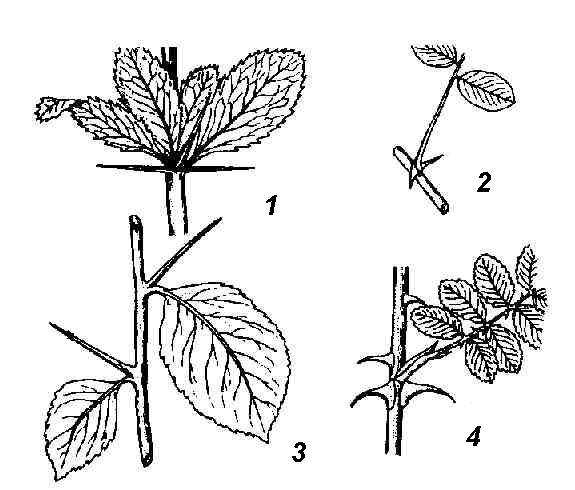
Rice. 4.26. Spines of various origins: 1 - barberry leaf spines; 2 - spines of white acacia, modification of stipules; 3 - spines of hawthorn shoot origin; 4 - thorns - rosehip emergents.
The shoots of a number of plants bear spikes. Thorns differ from spines in smaller sizes, these are outgrowths - emergents - of the integumentary tissue and tissues of the stem bark (rose hips, gooseberries) ( rice. 4.26, 4).
Adaptation to a lack of moisture is very often expressed in the early loss, metamorphosis or reduction of leaves that lose the main function of photosynthesis. This is compensated by the fact that the stem takes on the role of the assimilating organ. Sometimes such an assimilating stem of a leafless shoot remains externally unchanged (Spanish gorse, camel thorn) ( rice. 4.25, 3). The next step in this change of functions is the formation of such organs as phyllocladia and cladodia. These are flattened leaf-like stems or whole shoots. On the shoots of the needle ( rice. 4.25, 4), in the axils of scaly leaves, flat leaf-shaped phylloclades develop, which, like a leaf, have limited growth. Scale-like leaves and inflorescences are formed on phylloclades, which never happens on normal leaves, which means that the phyllocladium corresponds to a whole axillary shoot. Small, needle-like phylloclades are formed in asparagus in the axils of the scaly leaves of the main skeletal shoot. Cladodia are flattened stems that, unlike phyllocladia, retain the ability for long-term growth.
Some plants are characterized by the modification of leaves or their parts, and sometimes entire shoots in antennae, which twist around the support, helping the thin and weak stem to maintain an upright position. In many legumes, the upper part of the pinnate leaf (peas, peas, rank) turns into antennae. In other cases, stipules (sarsaparilla) turn into antennae. Very characteristic tendrils of leafy origin are formed in gourds, and all the transitions from normal to fully metamorphosed leaves can be seen. Antennae of shoot origin can be observed in grapes ( rice. 4.25, 2), passionflower and a number of other plants.