4.2. Escape and escape systems
The escape, like the root, is the main organ of the plant. Vegetative shoots typically perform the function of aerial nutrition, but have a number of other functions and are capable of various metamorphoses. spore-bearing shoots (including the flower) are specialized as organs reproductive providing reproduction.
The shoot is formed by the apical meristem as a whole and, therefore, is a single organ of the same rank as the root. However, compared with the root, the shoot has a more complex structure. The vegetative shoot consists of an axial part - stem, which is cylindrical in shape, and leaves- flat lateral organs sitting on the stem. In addition, an obligatory part of the escape are kidneys– rudiments of new shoots, which ensure the growth of the shoot and its branching, i.e. formation of the escape system. The main function of the shoot - photosynthesis - is carried out by leaves; stems are predominantly load-bearing organs that perform mechanical and conductive functions.
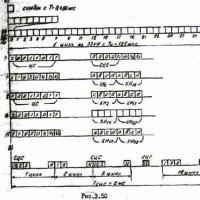
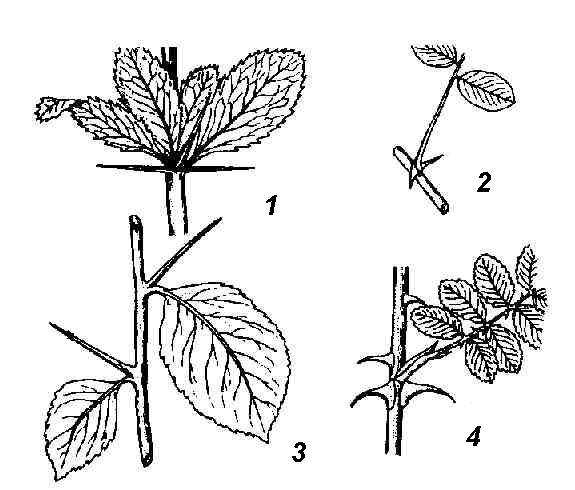
The main feature that distinguishes the shoot from the root is its foliage. The part of the stem from which the leaf (leaves) extends is called node. Stem segments between adjacent nodes internodes. Nodes and internodes are repeated along the axis of the shoot. So the escape has metameric structure, metamer(repeating element) of the shoot are the node with the leaf and the axillary bud and the underlying internode ( rice. 4.16).
Rice. 4.16. Escape structure.
The first shoot of a plant main escape, or escape of the first order. It is formed from an embryonic shoot ending kidney, which forms all subsequent metameres of the main shoot. By position, this kidney is apical; while it persists, this shoot is capable of further growth in length with the formation of new metameres. In addition to the apical, on the shoot are formed lateral kidneys. In seed plants, they are located in the axils of the leaves and are called axillary. From the lateral axillary buds develop lateral shoots, and branching occurs, due to which the total photosynthetic surface of the plant increases. Formed escape system, represented by the main shoot (shoot of the first order) and side shoots (shoots of the second order), and when branching is repeated, by side shoots of the third, fourth and subsequent orders. A shoot of any order has its own apical bud and is capable of growing in length.
Bud- this is a rudimentary, not yet unfolded shoot. Inside the kidney is the meristematic tip of the shoot - its apex(rice. 4.17). The apex is an actively working growth center that ensures the formation of all organs and primary tissues of the shoot. The source of constant self-renewal of the apex is the initial cells of the apical meristem, concentrated at the tip of the apex. The vegetative shoot apex, in contrast to the always smooth root apex, regularly forms protrusions on the surface, which are the beginnings of leaves. Only the very tip of the apex, which is called growth cone escape. Its shape varies greatly in different plants and does not always look like a cone; the apical part of the apex can be low, hemispherical, flat, or even concave.
From vegetative buds develop vegetative shoots consisting of a stem, leaves and buds. Such a kidney consists of a meristematic rudimentary axis ending growth cone, and rudimentary leaves of different ages. Due to uneven growth, the lower leaf primordia are bent inward and cover the upper, younger, leaf primordia and the growth cone. The nodes in the kidney are close together, since the internodes have not yet had time to stretch out. In the axils of leaf rudiments in the kidney, the rudiments of axillary buds of the following order can already be laid ( rice. 4.17). IN vegetative-generative a number of vegetative metameres are laid in the buds, and the growth cone is turned into a rudimentary flower or inflorescence. Generative, or floral the buds contain only the rudiment of an inflorescence or a single flower, in the latter case the bud is called bud.
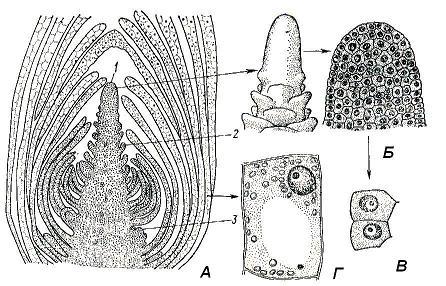
Rice. 4.17. The apical bud of the Elodea shoot: A - longitudinal section; B - growth cone (appearance and longitudinal section); C – cells of the apical meristem; D - parenchymal cell of the formed leaf; 1 - growth cone; 2 - leaf rudiment; 3 - the rudiment of the axillary kidney.
The outer leaves of the bud often change into kidney scales, which perform a protective function and protect the meristematic parts of the kidney from drying out and sudden changes in temperature. Such kidneys are called closed(wintering buds of trees and shrubs and some perennial grasses). open kidneys do not have kidney scales.
In addition to the usual, exogenous in inception, axillary buds, plants often form adnexal, or adventive kidneys. They arise not in the meristematic tip of the shoot, but on the adult, already differentiated part of the organ, endogenously, from internal tissues. Adnexal buds can form on stems (then they are usually located in internodes), leaves and roots. Adnexal buds are of great biological importance: they provide active vegetative renewal and reproduction of those perennial plants that have them. In particular, with the help of adnexal kidneys, they renew and multiply root offspring plants (raspberry, aspen, thistle, dandelion). Root offspring- these are shoots that have developed from adventitious buds on the roots. Adnexal buds on the leaves are formed relatively rarely. If such buds immediately give small shoots with adventitious roots that fall off the mother leaf and grow into new individuals, they are called brood(bryophyllum).
In the seasonal climate of the temperate zone, the deployment of shoots from the buds in most plants is periodic. In trees and shrubs, as well as in many perennial herbaceous plants, buds unfold into shoots once a year - in spring or early summer, after which new wintering buds are formed with the beginnings of next year's shoots. Shoots that grow from buds in one growing season are called annual shoots, or annual increments. In trees, they are well distinguished due to the formation renal rings- scars that remain on the stem after the fall of the kidney scales. In the summer of our deciduous trees, the annual shoots of only the current year are covered with leaves; there are no leaves on the annual shoots of previous years. In evergreen trees, leaves can be preserved on the corresponding annual increments of 3-5 past years. In a seasonally unseasoned climate, several shoots may form in one year, separated by small dormant periods. Such shoots formed in one growth cycle are called elementary shoots.
Buds that fall into a dormant state for a while, and then give new elementary and annual shoots, are called wintering or resting. According to their function, they can be called kidney regular renewal. Such buds are an obligatory feature of any perennial plant, woody or herbaceous, they ensure the perennial existence of an individual. By origin, renewal kidneys can be both exogenous (apical or axillary) and endogenous (adnexal).
If the lateral buds do not have a dormant period and develop simultaneously with the growth of the maternal shoot, they are called kidney enrichment. Deploying ones enrichment shoots greatly increase (enrich) the total photosynthetic surface of the plant, as well as the total number of inflorescences formed and, consequently, seed productivity. Enrichment shoots are typical for most annual herbs and for a number of perennial herbaceous plants with elongated flower-bearing shoots.
A special category is dormant buds, very characteristic of deciduous trees, shrubs, shrubs and a number of perennial grasses. By origin, they, like the buds of regular renewal, can be axillary and adnexal, but, unlike them, do not turn into shoots for many years. The stimulus for the awakening of dormant buds is usually either damage to the main trunk or branch (stump growth after cutting down a number of trees), or natural aging of the maternal shoot system associated with the attenuation of the vital activity of normal renewal buds (change of stems in shrubs). In some plants, leafless flowering shoots form from dormant buds on the trunk. This phenomenon is called caulifloria and is characteristic of many rainforest trees, such as the chocolate tree. In honey locust, bunches of large branched spines grow from sleeping buds on the trunk - modified shoots ( rice. 4.18).
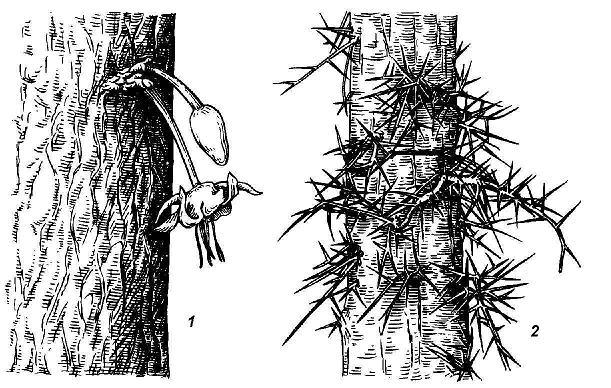
Rice. 4.18. Shoots from dormant buds: 1 - caulifloria near the chocolate tree; 2 - spines in honey locust from branched dormant buds.
Direction of shoot growth. Shoots growing vertically, perpendicular to the surface of the earth, are called orthotropic. Horizontally growing shoots are called plagiotropic. The direction of growth may change during shoot development.
Depending on the position in space, morphological types of shoots are distinguished ( rice. 4.19). The main shoot in most cases retains orthotropic growth and remains upright. Lateral shoots can grow in different directions, often forming a different angle with the parent shoot. In the process of growth, the shoot can change direction from plagiotropic to orthotropic, then it is called rising, or ascending. Shoots with plagiotropic growth that persists throughout life are called creeping. If they form adventitious roots at the nodes, they are called creeping.
Orthotropic growth is connected in a certain way with the degree of development of mechanical tissues. In the absence of well-developed mechanical tissues in elongated shoots, orthotropic growth is impossible. But often plants that do not have a sufficiently developed internal skeleton still grow upward. This is achieved in various ways. Weak shoots of such plants - creeper twist around some kind of solid support ( curly shoots), climb with the help of various kinds of spines, hooks, roots - trailers ( climbing shoots), cling with the help of antennae of various origins ( clinging shoots).
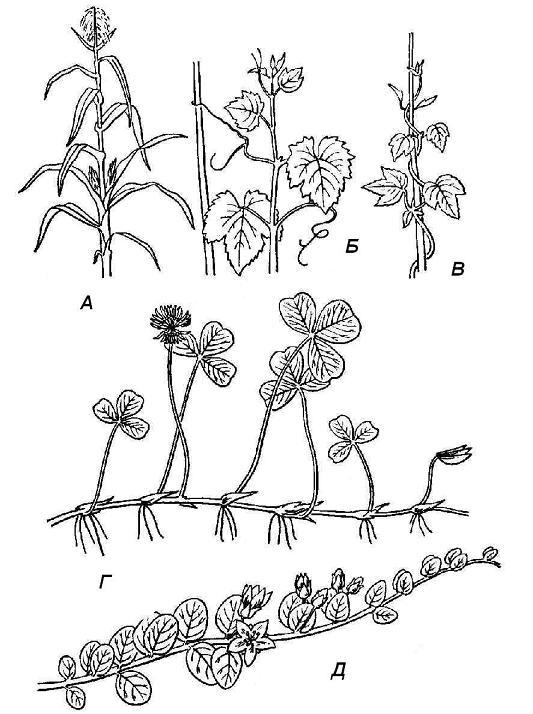
Rice. 4.19. Types of shoots by position in space: A - upright; B - clinging; B - curly; G - creeping; D - creeping.
Leaf arrangement. leaf arrangement, or phyllotaxis- the order of placement of leaves on the axis of the shoot. There are several main types of leaf arrangement ( rice. 4.20).
Spiral, or another leaf arrangement is observed when there is one leaf at each node, and the bases of successive leaves can be connected by a conditional spiral line. double row leaf arrangement can be considered as a special case of spiral. At the same time, at each node there is one sheet, covering the entire or almost the entire circumference of the axis with a wide base. Whorled leaf arrangement occurs when several leaves are laid on one node. Opposite leaf arrangement - a special case of whorled, when two leaves are formed on one node, exactly opposite each other; most often such a leaf arrangement occurs cross opposite, i.e. neighboring pairs of leaves are in mutually perpendicular planes ( rice. 4.20).
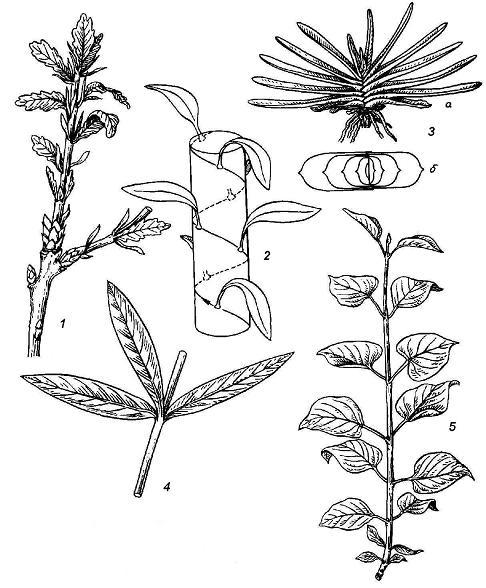
Rice. 4.20. Types of leaf arrangement: 1 - spiral in oak; 2 - scheme of spiral leaf arrangement; 3 - two-row in gasteria ( but- side view of the plant b– top view, scheme); 4 - whorled in oleander; 5 - opposite in lilac.
The order of initiation of leaf primordia on the shoot apex is a hereditary trait of each species, sometimes characteristic of a genus and even an entire family of plants. The leaf arrangement of the adult shoot is determined primarily by genetic factors. However, during the development of the shoot from the bud and its further growth, the location of the leaves can be influenced by external factors, mainly lighting conditions and gravity. Therefore, the final picture of leaf arrangement can differ greatly from the initial one and usually acquires a pronounced adaptive character. The leaves are arranged so that their plates are in the most favorable lighting conditions in each case. This is most pronounced in the form sheet mosaic observed on plagiotropic and rosette shoots of plants. In this case, the plates of all leaves are arranged horizontally, the leaves do not obscure each other, but form a single plane where there are no gaps; smaller leaves fill the gaps between the larger ones.
Shoot branching types. Branching is the formation of a system of axes. It provides an increase in the total area of contact of the plant body with air, water or soil. Branching arose in the process of evolution even before the appearance of organs. In the simplest case, the top of the main axis forks and gives rise to two axes of the next order. This apical, or dichotomous branching. Many multicellular algae have apical branching, as well as some primitive plants, such as club mosses ( rice. 4.21).
Other groups of plants are characterized by a more specialized side branch type. In this case, the lateral branches are laid below the top of the main axis, without affecting its ability to further increase. With this method, the potential for branching and formation of organ systems is much more extensive and biologically beneficial.
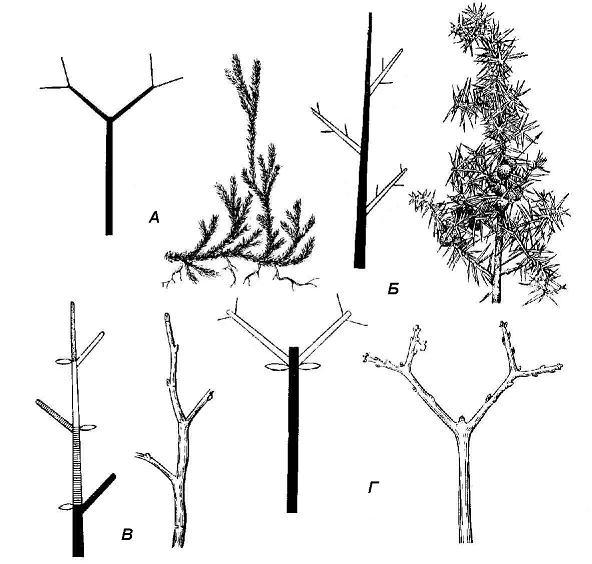
Rice. 4.21. Shoot branching types: A - dichotomous (club moss); B - monopodial (juniper); B - sympodial type of monochasia (bird cherry); D - sympodial according to the type of dichasia (maple).
There are two types of lateral branching: monopodial And sympodial(rice. 4.21). With a monopodial branching system, each axis is a monopodium, i.e. the result of the work of one apical meristem. Monopodial branching is characteristic of most gymnosperms and many herbaceous angiosperms. Most angiosperms, however, branch in a sympodial pattern. With sympodial branching, the apical bud of the shoot dies off at a certain stage or stops active growth, but an increased development of one or more lateral buds begins. Shoots are formed from them, replacing the shoot that has stopped growing. The resulting axis is a sympodium - a composite axis consisting of axes of several successive orders. The ability of plants to sympodial branching is of great biological importance. In case of damage to the apical bud, the growth of the axis will continue with lateral shoots.
Depending on the number of replacement axes, sympodial branching is distinguished by type monochasia, dichasia And pleiochasia. Branching according to the type of dichasia, or false dichotomous branching is typical for shoots with opposite leaf arrangement (lilac, viburnum).
In some groups of plants, the growth of the main skeletal axes occurs due to one or a few apical buds, lateral skeletal branches are not formed at all or are formed in a very small number. Tree-like plants of this type are found mainly in tropical areas (palm trees, dracaena, yucca, agave, cycads). The crown of these plants is formed not by branches, but by large leaves brought together in a rosette at the top of the trunk. The ability to rapidly grow and capture space, as well as to recover from damage in such plants is often absent or weakly expressed. Among temperate trees, such non-branching forms are practically not found.
The other extreme is plants that branch too profusely. They are represented by the life form cushion plants (rice. 4.22). The growth in the length of the shoots of these plants is extremely limited, but on the other hand, many lateral branches are formed annually, diverging in all directions. The surface of the shoot system of the plant looks as if trimmed; some pillows are so dense that they look like stones.
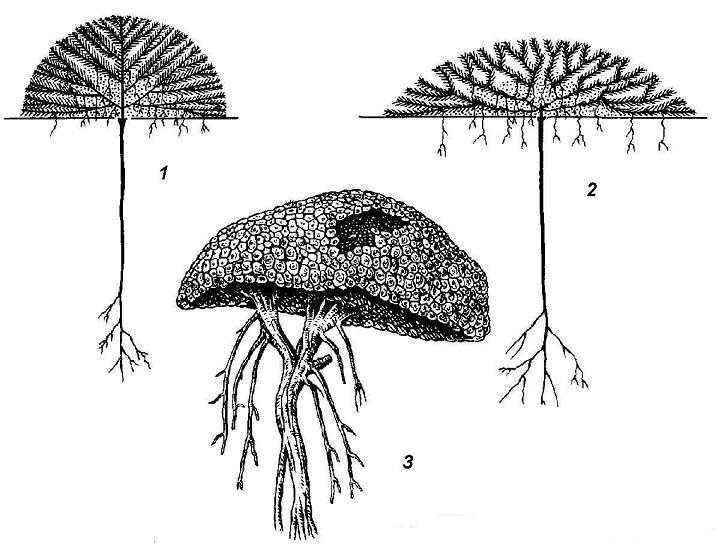
Rice. 4.22. Plants - pillows: 1, 2 - schemes of the structure of pillow plants; 3 - Azorella from Kerguelen Island.
Representatives of a life form branch very strongly Tumbleweed characteristic of steppe plants. A spherically branched, very loose system of shoots is a huge inflorescence, which, after fruit ripening, breaks off at the base of the stem and rolls over the steppe with the wind, scattering the seeds.
Specialization and metamorphoses of shoots. Many plants within the shoot system have a certain specialization. Orthotropic and plagiotropic, elongated and shortened shoots perform different functions.
elongated called shoots with normally developed internodes. In woody plants, they are called growth and are located along the periphery of the crown, determining its shape. Their main function is to capture space, increase the volume of photosynthetic organs. shortened shoots have close nodes and very short internodes ( rice. 4.23). They form inside the crown and absorb scattered light penetrating there. Often shortened shoots of trees are flowering and perform the function of reproduction.
Rice. 4.23. Shortened (A) and elongated (B) sycamore shoots: 1 - internode; 2 - annual increments.
Herbaceous plants usually have shortened rosette shoots perform the function of perennial skeletal and photosynthetic, and elongated ones are formed in the axils of rosette leaves and are flower-bearing (plantain, cuff, violets). If axillary peduncles are leafless, they are called arrows. The fact that flowering shoots are short in woody plants and elongated in herbaceous plants is biologically well explained. For successful pollination, grass inflorescences must be raised above the herbage, and in trees, even shortened shoots in the crown are in favorable conditions for pollination.
An example of the specialization of shoots is the perennial axial organs of woody plants - trunks And branches crowns. In deciduous trees, annual shoots lose their assimilation function after the first growing season, in evergreen trees - after a few years. Some of the shoots die off completely after the loss of leaves, but the majority remain as skeletal axes, performing supporting, conducting, and storage functions for decades. The leafless skeletal axes are known as boughs And trunks(by the trees) stems(for shrubs).
In the course of adaptation to specific environmental conditions or in connection with a sharp change in functions, shoots can change (metamorphize). Shoots developing underground are especially often metamorphosed. Such shoots lose the function of photosynthesis; they are common in perennial plants, where they act as organs for experiencing an unfavorable period of the year, stock and renewal.
The most common underground shoot metamorphosis is rhizome (rice. 4.24). It is customary to call a rhizome a long-lived underground shoot that performs the functions of deposition of reserve nutrients, renewal, and sometimes vegetative propagation. The rhizome is formed in perennial plants, which, as a rule, do not have a main root in the adult state. According to its position in space, it can be horizontal, oblique or vertical. The rhizome usually does not bear green leaves, but, being a shoot, retains a metameric structure. The nodes are distinguished either by leaf scars and the remains of dry leaves, or by living scaly leaves, and axillary buds are also located in the nodes. According to these features, the rhizome is easy to distinguish from the root. As a rule, adventitious roots are formed on the rhizome; lateral branches of the rhizome and above-ground shoots grow from the buds.
The rhizome is formed either initially as an underground organ (kupena, raven eye, lily of the valley, blueberry), or first as an above-ground assimilating shoot, which then sinks into the soil with the help of retracting roots (strawberry, lungwort, cuff). Rhizomes can grow and branch monopodially (cuff, crow's eye) or sympodially (kupena, lungwort). Depending on the length of the internodes and the intensity of growth, there are long And short rhizomes and, accordingly, long-rhizome And short-rhizome plants.
When branching rhizomes, it is formed curtain elevated shoots connected by sections of the rhizome system. If the connecting parts are destroyed, the shoots are isolated, and vegetative reproduction occurs. The totality of new individuals formed vegetatively is called clone. Rhizomes are characteristic mainly of herbaceous perennials, but are also found in shrubs (euonymus) and shrubs (lingonberries, blueberries).
close to roots underground stolons- short-lived thin underground shoots bearing underdeveloped scaly leaves. Stolons serve for vegetative reproduction, settlement and territory capture. Spare nutrients are not deposited in them.
In some plants (potato, earth pear), by the end of summer, stolons form from the apical buds of stolons. tubers (Fig. 4.24). The tuber has a spherical or oval shape, the stem is strongly thickened, reserve nutrients are deposited in it, the leaves are reduced, and buds form in their axils. The stolons die off and collapse, the tubers overwinter, and the next year they give rise to new above-ground shoots.
Tubers do not always develop on stolons. In some perennial plants, the base of the main shoot grows tuberous and thickens (cyclamen, kohlrabi cabbage) ( rice. 4.24). The functions of the tuber are a supply of nutrients, experiencing an unfavorable period of the year, vegetative renewal and reproduction.
In perennial grasses and dwarf shrubs with a well-developed tap root that persists throughout life, a kind of organ of shoot origin is formed, called caudex. Together with the root, it serves as a place for the deposition of reserve substances and carries many renewal buds, some of which may be dormant. The caudex is usually subterranean and is formed from short shoot bases that sink into the soil. Caudex differs from short rhizomes in the way it dies off. Rhizomes, growing at the top, gradually die off and collapse at the older end; the main root is not preserved. The caudex grows in width, from the lower end it gradually turns into a long-lived thickening root. The death and destruction of the caudex and the root goes from the center to the periphery. A cavity is formed in the center, and then it can be divided longitudinally into separate sections - particles. The process of dividing an individual of a taproot plant with a caudex into parts is called particulation. There are many caudex plants among legumes (lupins, alfalfa), umbrella plants (femur, ferula), and Compositae (dandelion, wormwood).
Bulb- this is usually an underground shoot with a very short flattened stem - bottom and scaly fleshy succulent leaves that store water and soluble nutrients, mainly sugars. Aerial shoots grow from the apical and axillary buds of the bulbs, adventitious roots form on the bottom ( rice. 4.24). Thus, the bulb is a typical organ of vegetative renewal and reproduction. Bulbs are most characteristic of plants from the families of lilies (lilies, tulips), onions (onions) and amaryllis (daffodils, hyacinths).
The structure of the bulb is very diverse. In some cases, bulbs storing scales are only modified leaves that do not have green plates (lily saranka); in others, these are underground sheaths of green assimilating leaves, which thicken and remain in the bulb after the plates die (onion). Bulb axis growth can be monopodial (snowdrop) or sympodial (hyacinth). The outer scales of the bulb consume the supply of nutrients, dry out and play a protective role. The number of onion scales varies from one (garlic) to several hundred (lilies).
As an organ of renewal and reserve, the bulb is adapted mainly to climates of the Mediterranean type - with fairly mild, wet winters and very hot, dry summers. It serves not so much for a safe overwintering, but for experiencing a harsh summer drought. The storage of water in the tissues of onion scales occurs due to the formation of mucus, which can retain a large amount of water.
Corm outwardly resembles an onion, but its scaly leaves are not storage; they are dry and membranous, and reserve substances are deposited in the thickened stem part (saffron, gladiolus).

Rice. 4.24. Underground escape metamorphoses: 1, 2, 3, 4 - sequence of development and structure of the potato tuber; 5 - cyclamen tuber; 6 - kohlrabi tuber; 7 - bulbs of a tiger lily; 8 - onion bulb; 9 - lily bulb; 10 - section of a long rhizome of couch grass.
Not only underground, but also above-ground shoots of plants can be modified ( rice. 4.25). Quite common elevated stolons. These are plagiotropic short-lived shoots, the function of which is vegetative reproduction, resettlement and territory capture. If stolons carry green leaves and participate in the process of photosynthesis, they are called lashes(bone, tenacious creeping). In strawberries, stolons are devoid of developed green leaves, their stems are thin and fragile, with very long internodes. Such more highly specialized stolons for the function of vegetative reproduction are called mustache.
Juicy, fleshy, adapted to the accumulation of water can be not only bulbs, but also above-ground shoots, usually in plants living in conditions of lack of moisture. Water storage organs can be leaves or stems, sometimes even buds. Such succulent plants are called succulents. Leaf succulents store water in leaf tissues (aloe, agave, jughead, rhodiola, or golden root). Stem succulents are characteristic of the American cactus family and African euphorbiaceae. The succulent stem performs a water-reserving and assimilating function; leaves are reduced or turned into spines ( rice. 4.25, 1). In most cacti, the stems are columnar or spherical, leaves are not formed on them at all, but the nodes are clearly visible by the location of the axillary shoots - areola having the appearance of warts or elongated outgrowths with spines or tufts of hairs. The transformation of leaves into spines reduces the evaporative surface of the plant and protects it from being eaten by animals. An example of the metamorphosis of a kidney into a succulent organ is head of cabbage serves as a cultivated cabbage.
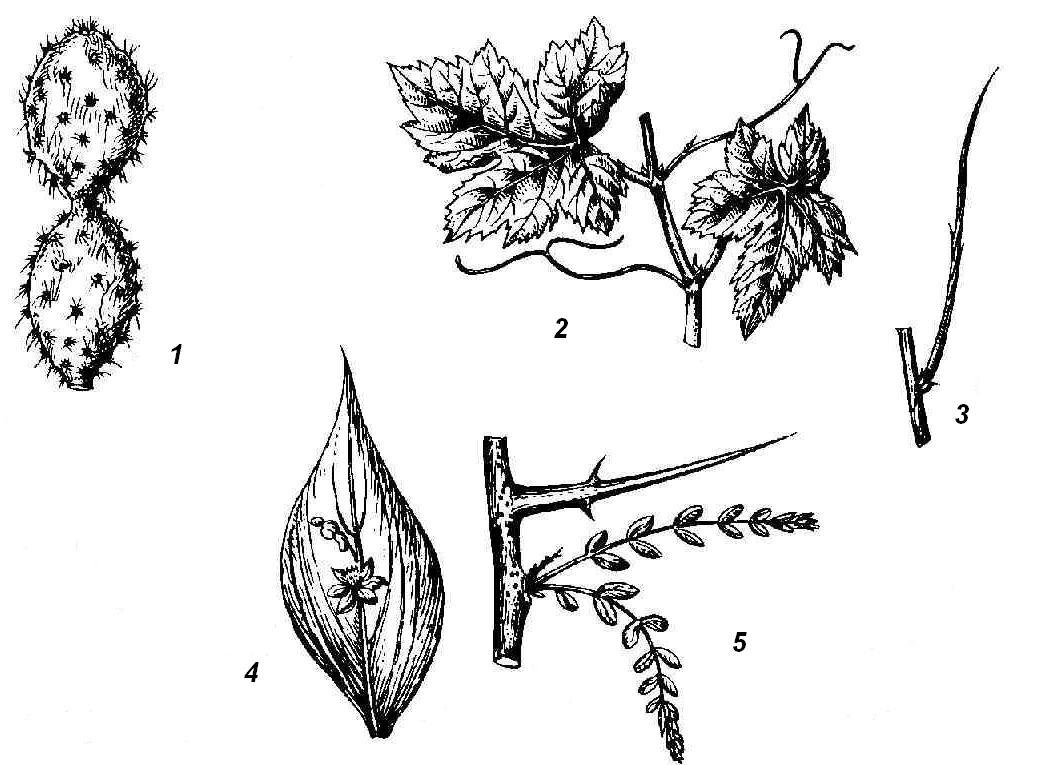
Rice. 4.25. Elevated shoot metamorphoses: 1 - stem succulent (cactus); 2 - tendrils of grapes; 3 - leafless photosynthetic shoot of gorse; 4 - phyllocladium of butcher's broom; 5 - thorn of honey locust.
spines cacti are leafy. Leaf spines are often found in non-succulent plants (barberry) ( rice. 4.26, 1). In many plants, spines are not of leaf, but of stem origin. In the wild apple tree, wild pear, laxative joster, shortened shoots metamorphosed into spines, having limited growth and ending in a point. They acquire the appearance of a hard lignified thorn after the leaves fall. At the hawthorn ( rice. 4.26, 3) the spines that form in the axils of the leaves are completely leafless from the very beginning. In honey locust ( rice. 4.25.5) powerful branched spines are formed on trunks from dormant buds. The formation of spines of any origin, as a rule, is the result of a lack of moisture. When many thorny plants are grown in an artificial humid atmosphere, they lose their spines and instead grow normal leaves (camel thorn) or leafy shoots (English gorse).
Rice. 4.26. Spines of various origins: 1 - barberry leaf spines; 2 - spines of white acacia, modification of stipules; 3 - spines of hawthorn shoot origin; 4 - thorns - rosehip emergents.
The shoots of a number of plants bear spikes. Thorns differ from spines in smaller sizes, these are outgrowths - emergents - of the integumentary tissue and tissues of the stem bark (rose hips, gooseberries) ( rice. 4.26, 4).
Adaptation to a lack of moisture is very often expressed in the early loss, metamorphosis or reduction of leaves that lose the main function of photosynthesis. This is compensated by the fact that the stem takes on the role of the assimilating organ. Sometimes such an assimilating stem of a leafless shoot remains externally unchanged (Spanish gorse, camel thorn) ( rice. 4.25, 3). The next step in this change of functions is the formation of such organs as phyllocladia And cladodia. These are flattened leaf-like stems or whole shoots. On the shoots of the needle ( rice. 4.25, 4), in the axils of scaly leaves, flat leaf-shaped phylloclades develop, which, like a leaf, have limited growth. Scale-like leaves and inflorescences are formed on phylloclades, which never happens on normal leaves, which means that the phyllocladium corresponds to a whole axillary shoot. Small, needle-like phylloclades are formed in asparagus in the axils of the scaly leaves of the main skeletal shoot. Cladodia are flattened stems that, unlike phyllocladia, retain the ability for long-term growth.
Some plants are characterized by the modification of leaves or their parts, and sometimes entire shoots in antennae, which twist around the support, helping the thin and weak stem to maintain an upright position. In many legumes, the upper part of the pinnate leaf (peas, peas, rank) turns into antennae. In other cases, stipules (sarsaparilla) turn into antennae. Very characteristic tendrils of leafy origin are formed in gourds, and all the transitions from normal to fully metamorphosed leaves can be seen. Antennae of shoot origin can be observed in grapes ( rice. 4.25, 2), passionflower and a number of other plants.