Жгутики, ворсинки, пили. Способы передвижения прокариот. Методы определения подвижности у бактерий. Определение подвижности микроорганизмов Методы изучения подвижности бактерий
Для эффективного воздействия на бактериальный состав среды необходимо иметь достоверную информацию о его качественном и количественном содержании. Существует много методов определения бактерий, и выбор способа изучения образца зависит от того, какие результаты должны быть получены. Метод определения молочнокислых бактерий отличается от методики выявления Listeria monocytogenes, а определение ферментативной активности производится иначе, чем определение биохимических свойств.
Методы определения по получаемому результату можно разделить на две большие группы:
- определение количества микробов;
- качественное исследование.
Результаты анализа бактериального состава образцов выражают общим микробным числом, выраженным в КОЕ (колониеобразующие единицы).
Анализ количества микробов, в зависимости от его возможностей, может определять:
- число всех микроорганизмов, содержащихся в образце;
- только жизнеспособные микробы.
В зависимости от способа получения результата, методы определения количества микроорганизмов подразделяют на:
- прямые (микроскопические);
- косвенные.
В свою очередь, косвенные методы разделяют в зависимости от применяемого критерия как:
- методы оптического исследования (спектрофотометрия, нефелометрия) – измеряемый параметр зависит от количества микроорганизмов;
- высев – метод измерения образовавшихся колоний.

Методы определения общего количества бактерий опираются на значение титра образца.
Методика титра
Метод предельного разбавления образца (метод титра) позволяет с высокой точностью определить количественное значение группы микроорганизмов.
Сущность методики заключается в том, что исследуемая проба разводится определенным образом и высевается в специфические для микроорганизмов среды. Так создаются благоприятные условия для роста. По прошествии времени исследуют образцы, устанавливая, при каком предельном разведении выявляются бактерии определенной группы. Выводы делаются по специфическим изменениям питательного субстрата.

Подобная методика, учитывающая индивидуальные свойства микробов, хорошо зарекомендовала себя при обнаружении микроорганизмов кишечной палочки и родственных ей видов.
Прямой подсчет
Метод удобен для исследования проб почвы и воды. Осуществляется прямой подсчет в предназначенных для этого счетных камерах, на мембранных фильтрах или фиксированных мазках. Метод не требует сложного оборудования, непродолжителен по времени и минимален по стоимости.
Ограничением использования метода является обязательная высокая концентрация микробов в образцах.
Метод оптического определения количества бактерий, основанный на определении светорассеивания взвесью образца. Данный способ позволяет определить число клеток в образце, что делает метод востребованным при микробиологических исследованиях.

Метод нефелометрии
Подсчет жизнеспособных микробов
Методика базируется на посеве определенного количества бактерий в виде суспензии на среду агара. После этого осуществляют подсчет сформированных колоний, имея в виду, что каждая из них является потомством жизнеспособной бактерии.
Существует две разновидности способа посева образца:
- исследуемый образец вносится в среду агара и перемешивается;
- образец высевается на поверхностном слое агара.
Подвижность как важный фактор идентификации бактерий
Значимым фактором идентификации бактерий является подвижность, которая обеспечивается жгутиками. Так как количество и расположение жгутиков, обеспечивающих подвижность, может быть различным, все имеющие жгутики микробы подразделяют для удобства идентификации на:
- монотрихи – один жгутик на полюсе;
- лофотрихи – пучок жгутиков, расположенный на одном из полюсов;
- амфитрихи – жгутики или пучки расположены на обоих полюсах;
- перитихи – жгутики расположены по периметру клетки.

Определение подвижности бактерий проводят в культурах не старше суток. У старых культур способность передвигаться утрачивается.
Определение качественного бактериального состава
Определение качественного состава бактерий опирается на несколько факторов.
Биолого-химические особенности микроорганизмов
Изучение биохимических свойств микроорганизмов помогает определению качественного состава бактерий.
Идентификации микроорганизмов способствует знание биохимических процессов. Методы определения количества и качества бактерий опираются на протеолитические и сахаролитические свойства микроорганизмов, а также токсино- и пигментообразование.
Ферменты микробов
Одним из факторов жизнедеятельности бактерий является ферментативная активность: ферментный состав и свойства регламентируются геномом микроорганизма и является стабильным критерием идентификации микробов. Поэтому обнаружение протеолитических, сахаролитических и других ферментов имеет большое значение в идентификации микроорганизмов.

К примеру, критерием протеолитической активности микроорганизмов является способность бактерий расщеплять белок до продуктов глубокого распада (сероводород и индол). На этом результате ферментативной активности основан метод определения числа микроорганизмов, имеющий важное практическое значение.
Свойство пигментообразования
Другим стойким генетическим признаком бактерий является пигментообразование. Данное свойство предназначено для защиты бактериальной клетки от воздействия ультрафиолетовых лучей.
Большая часть патогенных микроорганизмов не обладает подобными защитными свойствами – пигментообразование для них не характерно.
Изучение микрофлоры молока
Определение бактерий имеет значение в практической деятельности человека, например, в пищевой промышленности. Так, бактериальная обсемененность молока является основным показателем санитарных условий его получения. В случае превышения порогового количества микроорганизмов в молоке, сортность продукта снижается.
С 1987 г. страны ЕЭС приняли единые стандарты по степени бактериальной обсемененности молока, подразделяя продукт на три категории:
- А – 20 тыс./мл;
- В – 100 тыс./мл;
- С – свыше 100 тыс./мл.

В данном случае числа указывают на максимально возможное количество микроорганизмов в 1 мл молока (обсемененность).
Бактериальная обсемененность молока напрямую зависит от санитарных условий получения и первоначальной обработки продукта. Так, использование для очистки молока многоразовых фильтров может приводить к дополнительному бактериальному обсеменению.
Наличие в молоке соматических клеток является важным критерием качества. Эти клетки являются частичками биомассы животного. Они образуются в вымени и отражают естественные процессы старения и обновления организма.
Число соматических клеток в молоке возрастает при наличии у животных травм, заболеваний ЖКТ или других патологий, что приводит к росту показателя бактериальной обсемененности молока.
Определение молочнокислых бактерий
Большое значение в пищевой промышленности уделяется молочнокислым микроорганизмам благодаря их антагонистической и протеолитеческой активности.
К примеру, пластичная консистенция и выраженный вкус различных сортов сыра связаны с протеолитической активностью молочнокислых микробов закваски. Исследованиям активностью молочнокислых бактерий в этой области уделяется большое значение, но, до сих пор не удалось создать критерий определения штаммов молочнокислых бактерий закваски по показателю протеолитеческой активности.

Антагонистическая активность молочнокислых микробов используется не только в пищевой промышленности, но и в медицине, ветеринарии сельском хозяйстве и т.д.
Примеры применения молочнокислых бактерий как антагонистов к определенным микроорганизмам:
- производство сыров – антагонисты к масляным микробам и кишечной палочке;
- хлебопекарное производство – антагонисты к споровой палочке, возбудителю «картофельной болезни» хлеба;
- молочнокислые продукты – антагонисты к бактериям, провоцирующим развитие желудочно-кишечных инфекций.
При необходимости подсчета в закваске количества молочнокислых бактерий используют метод посева микробов на агаре и молоке с добавлением мела. Образующаяся молочная кислота растворяет мел, и вокруг колоний молочнокислых бактерий появляются светлые зоны.
Для подсчета молочнокислых стрептококков используют метод предельного разведения, высевая их в молоко. В зависимости от термофильности молочнокислых бактерий выбирается оптимальный термальный режим посева. Образцы со свернувшимся молоком используют для приготовления микроскопических препаратов. После этого выявляют минимальное разведение, содержащее молочнокислые палочки.
Воздушный микробиологический контроль
Для жизнедеятельности бактерий воздушная среда не является благоприятной, но большинство микроорганизмов, попадая в воздух, способны временно сохранять активность и свои свойства. Среди них такие патогенные бактерии, как возбудители кори, скарлатины, коклюша, оспы, туберкулеза, легочной чумы и другие инфекции дыхательных путей, передающиеся воздушно-капельным методом.

Микробиологический контроль воздушной среды оценивает общую бактериальную обсемененность воздуха и разрабатывает профилактические методы снижения числа возбудителей инфекционных заболеваний.
Объектами исследования степени бактериальной обсемененности в закрытых помещениях являются больницы и поликлиники, детские учреждения и места постоянного скопления людей (кинотеатры, спортзалы и другие). Определение степени бактериальной обсемененности воздуха в помещениях проводится по отработанным методикам, включающим следующие действия:
- забор образца;
- транспортировка и подготовка пробы;
- бактериальный посев;
- определение микроорганизмов посредством идентификации.
В случае выявления высокой степени бактериальной обсемененности, для понижения количества бактерий используют различные методы:
- химические – обработка помещения двуокисью азота, озоном или суспензией молочной кислоты;
- механические – принудительная фильтрация воздуха;
- физические – облучение ультрафиолетом.
Исследование бактериального состава воды
Анализ воды проводят на наличие следующих групп микроорганизмов:
- колиформные микробы – микроорганизмы группы кишечной палочки, используются как маркеры фекальной контаминации (обсеменения);
- клостридии – микробы, обладающие высокой устойчивостью к обеззараживаннию; реперный показатель (ориентир) – если в пробе нет клостридий, то нет и других патогенных микробов;
- вирусы;
- лямблии.
Колиформные бактерии являются грамотрицательными бактериями группы кишечной палочки, обитающими в кишечнике млекопитающих и птиц. В воду они попадают с фекалиями, способны существовать в ней неделями, но теряют свойство размножения.
Наличие колиформных микроорганизмов в образце воды указывает на высокую вероятность присутствия сточных вод. Наличие вирулентных (болезнетворных) штаммов колиформных бактерий – показатель риска возникновения заболеваний.
Колиформные микробы включают в себя группу термотолерантных колиформных микроорганизмов, обладающих свойством выживать при высоких (45°С) температурах. Термотолерантные колиформные бактерии являются индикатором недавнего фекального заражения, аналитически легко определяются.
Согласно СанПиН, колиформные бактерии не могут присутствовать в системах водоснабжения, а наличие колиформных микроорганизмов указывает либо на некачественную очистку, либо на вторичное фекальное загрязнение. Нормой считается наличие колиформных микробов в количестве не более 5% от общего числа. Критерием эффективности очистки от фекальных стоков являются термотолерантные колиформные бактерии, как легко определяемые.
Выявление возбудителя листериоза
Бактерия Listeria monocytogenes является возбудителем листериоза – инфекционного заболевания людей и животных.

Listeria monocytogenes
Возбудитель листериоза – имеющая высокую подвижность неспорообразующая грамположительная кишечная палочка Listeria monocytogenes. У людей заражение Listeria monocytogenes протекает как острый сепсис, поражая центральную нервную систему, лимфосистему, миндалины, селезенку и печень. Поражение Listeria monocytogenes у человека может протекать как в острой, так и в хронической форме.
Статистическая группа риска по Listeria monocytogenes:
- люди преклонного возраста;
- беременные;
- лица с ослабленным иммунитетом, ВИЧ-инфицированные и онкобольные;
- алкоголики и наркоманы.
Определяется зараженность Listeria monocytogenes методом ПЦР (полимеразная цепная реакция) – в плазме крови выявляется ДНК Listeria monocytogenes. Подтверждением наличия листериоза является выявление специфичного участка ДНК Listeria monocytogenes. Специфичность определения ДНК Listeria monocytogenes – 100%; метод является высокочувствительным.
Взаимодействие бактерий с антибиотиками
Выявление чувствительности бактерий к антибиотикам имеет большое практическое значение для расчета дозировки препаратов при профилактике и лечении инфекционных заболеваний.

Для выявления чувствительности микробов к различным антибиотикам применяются два основных метода:
- определение чувствительности к антибиотиками при помощи дисков;
- изучение чувствительности микроорганизмов к антибиотикам посредством серийного разведения.
Методика с использованием дисков
Для выявления степени чувствительности к антибиотикам, в питательную культуру засевают изучаемые бактерии. На поверхности размещают диски, содержащие различные антибиотики в известных дозировках.
Пробу выдерживают при оптимальной температуре (37°С) сутки, после чего, сравнивая диаметр кольцевых безмикробных зон вокруг различных дисков, делают вывод о чувствительности бактерий к различным антибиотикам и их концентрациям.
Для получения воспроизводимых результатов следует пользоваться стандартными дисками и питательной средой, а в качестве контроля применять эталонные штаммы. Методика дисков не позволяет получить надежные результаты, а так же очень критична к слабодифундирующим антибиотикам (ристомицин или полимиксин).
Серийное разведение
Чувствительность микробов к антибиотикам методом серийного разведения позволяет выявить минимальную концентрацию лекарственного препарата, оказывающую терапевтический эффект. Чувствительность микроорганизмов определяют, как:
- чувствительные штаммы – жизнедеятельность бактерий подавляется обычной дозировкой антибиотиков в крови;
- умеренно устойчивые штаммы – для ингибирования бактерий необходимо использование максимальных доз антибиотиков;
- устойчивые бактерии – жизнедеятельность бактерий не подавляется даже при максимальной концентрации антибиотика, то есть отсутствие чувствительности.

Определение чувствительности бактериофагов
Бактериофаги являются естественными врагами бактерий. Характер взаимодействия бактериофага с бактериями описывается как:
- вирулентное, вызывающее лизис (гибель) микробной клетки;
- умеренное – переход бактерии в неинфекционную форму фага (профаг).
Бактериофаги специфичны к определенным группам микроорганизмов, что отражается в их названии – стрептококковые фаги, стафилококковые фаги и т.д. Способом определения количества бактериофагов в единице объема является метод агаровых слоев. Он прост в исполнении и обладает достаточной точностью.
Таким образом, существует большое количество методов определения микроорганизмов. Выбор оптимального зависит от заданного критерия отбора.

Работаю врачом ветеринарной медицины. Увлекаюсь бальными танцами, спортом и йогой. В приоритет ставлю личностное развитие и освоение духовных практик. Любимые темы: ветеринария, биология, строительство, ремонт, путешествия. Табу: юриспруденция, политика, IT-технологии и компьютерные игры.
Подвижность бактерий может обеспечиваться различным образом. У большинства активно передвигающихся, плавающих бактерий движение обусловлено вращением жгутиков. Двигаться без жгутиков способны скользящие бактерии (к которым относятся миксобактерии, цианобактерии и некоторые другие группы) и спирохеты. О механизмах их движения будет сказано при рассмотрении соответствующих групп бактерий. Расположение жгутиков.Расположение жгутиков у подвижных эубактерий - это признак, характерный для определенных групп, поэтому оно имеет таксономическое значение. У палочковидных бактерий жгутики могут прикрепляться полярноили латерально(рис. 2.34). Среди бактерий с монополярным жгутикованием лишь немногие снабжены только одним, но зато особенно толстым жгутиком - это монотрихи (Vibrio metschnikovii, рис. 2.35; Caulobacter sp.). У многих бактерий с монополярным и биполярным жгутикованием одиночный по виду жгутик в действительности представляет собой пучок из 2-50 жгутиков (политрихи). Монополярно-политрихальное расположение жгутиков называют также лофотрихальным(как у Pseudomonas, Chromatium), а биполярно-политрихальное - амфитрихальным (у Spirillum). У Selenomonas имеется один пучок жгутиков, прикрепленный сбоку (рис. 2.36,2>). При перитрихальномрасположении (как у Enterobacteriaceae, Bacillaceae и не которых других бактерий) жгутики располагаются по бокам клетки или на всей поверхности (рис. 2.36,4).

 Выявление жгутиков.Рассмотреть жгутик (или пучок жгутиков) в проходящем свете или в условиях фазового контраста удается только у немногих бактерий, например у Chromatium okenii, Bdellovibrio,Thiospirillum
(рис. 2.37). У многих других бактерий (Pseudomonas, Spirillum
и др.) жгутик и зону его биения можно увидеть только в темном поле. Легче всего выявлять жгутики путем нанесения на них красителя или металла, а также с помощью электронного микроскопа.
Функции жгутиков.У большинства бактерий с полярным расположением жгутиков последние действуют подобно корабельному винту и проталкивают клетку в окружающей жидкой среде. Жгутик представляет собой спирально извитую нить, приводимую во вращательное движение «мотором», находящимся в месте ее прикрепления в плазматической мембране. Для перемещения клетки может служить одиночный жгутик или пучок жгутиков. Жгутики вращаются сравнительно быстро; например, у спирилл они совершают около 3000 оборотов в минуту, что близко к скорости среднего электромотора. Вращение жгутиков приводит к тому, что тело клетки вращается примерно с 1/3 этой скорости в противоположном направлении.
Жгутики могут спонтанно или в ответ на внешний стимул изменять направление вращения (рис. 2.34). У некоторых бактерий с полярным расположением жгутиков это приводит к тому, что клетка начинает двигаться вспять. Когда у Chromatium okenii
в ответ на вспышку света направление вращения жгутиков меняется, пучок жгутиков превращается в тянущее приспособление; при этом назад клетка перемещается в четыре раза медленнее, чем вперед, и ее движение становится «кувыркающимся». У Thiospirillum jenense -
гигантской фототрофной спириллы - единственный полярный пучок жгутиков при обратном движении бьется уже не впереди клетки: пространство биения жгутиков теперь охватывает клетку с боков: оно как бы вывернуто наизнанку (подобно вывернутому ветром зонту). У спирилл с амфитрихальным расположением жгутиков в таком положении находится, смотря по обстоятельствам, то один, то другой пучок.
Перитрихально расположенные жгутики Escherichia coli
работают как один хорошо скоординированный спиральный пучок и проталкивают клетку через среду. В тех случаях, когда направление вращения отдельных жгутиков меняется, клетка начинает «кувыркаться». По-видимому, перитрихально расположенные жгутики не могут служить тянущим приспособлением.
Бактерии, снабженные жгутиками, могут передвигаться очень быстро: Bacillus megaterium
со скоростью 1,6 мм/мин, Vibrio cholerae -
12 мм/мин. Это соответствует примерно от 300 до 3000 длин тела в минуту.
Тонкое строение жгутиков.Жгутики представляют собой спирально закрученные нити. У разных бактерий они различаются по своей толщине (12-18 нм), длине (до 20 мкм), а также по длине и амплитуде витка. Эти параметры характерны для каждого вида. У некоторых бактерий могут образовываться жгутики разных типов. Нити жгутиков состоят из специфического белка флагеллина. Они построены из субъединиц с относительно малой молекулярной массой. Субьединицы располагаются по спирали вокруг внутреннего свободного пространства (подобно белковым молекулам в вирусе табачной мозаики). Таким образом, структура жгутика определяется свойствами белковых субъединиц.
Жгутик состоит из трех частей - описанной выше спиральной нити, «крюка» вблизи поверхности клетки и базального тельца. С помощью базального тельца жгутик закреплен в плазматической мембране и в клеточной стенке (рис. 2.38). Оно состоит из центрального стержня, на котором у грам-отрицательных бактерий находятся две пары колец. Наружная пара (кольца L и Р) расположены на уровне наружного и внутреннего слоев клеточной стенки, а внутренняя пара (кольца S и М) - на уровне наружного слоя плазматической мембраны. Так как у грам-положительных бактерий наружная пара колец отсутствует, полагают, что для вращения жгутиков необходима только внутренняя пара. Можно представить себе, что кольцо М действует как приводной диск, а кольцо S играет роль подшипника на внутренней поверхности пептидогликанового слоя. Молекулярный механизм вращательного «мотора» жгутика пока не выяснен.
Выявление жгутиков.Рассмотреть жгутик (или пучок жгутиков) в проходящем свете или в условиях фазового контраста удается только у немногих бактерий, например у Chromatium okenii, Bdellovibrio,Thiospirillum
(рис. 2.37). У многих других бактерий (Pseudomonas, Spirillum
и др.) жгутик и зону его биения можно увидеть только в темном поле. Легче всего выявлять жгутики путем нанесения на них красителя или металла, а также с помощью электронного микроскопа.
Функции жгутиков.У большинства бактерий с полярным расположением жгутиков последние действуют подобно корабельному винту и проталкивают клетку в окружающей жидкой среде. Жгутик представляет собой спирально извитую нить, приводимую во вращательное движение «мотором», находящимся в месте ее прикрепления в плазматической мембране. Для перемещения клетки может служить одиночный жгутик или пучок жгутиков. Жгутики вращаются сравнительно быстро; например, у спирилл они совершают около 3000 оборотов в минуту, что близко к скорости среднего электромотора. Вращение жгутиков приводит к тому, что тело клетки вращается примерно с 1/3 этой скорости в противоположном направлении.
Жгутики могут спонтанно или в ответ на внешний стимул изменять направление вращения (рис. 2.34). У некоторых бактерий с полярным расположением жгутиков это приводит к тому, что клетка начинает двигаться вспять. Когда у Chromatium okenii
в ответ на вспышку света направление вращения жгутиков меняется, пучок жгутиков превращается в тянущее приспособление; при этом назад клетка перемещается в четыре раза медленнее, чем вперед, и ее движение становится «кувыркающимся». У Thiospirillum jenense -
гигантской фототрофной спириллы - единственный полярный пучок жгутиков при обратном движении бьется уже не впереди клетки: пространство биения жгутиков теперь охватывает клетку с боков: оно как бы вывернуто наизнанку (подобно вывернутому ветром зонту). У спирилл с амфитрихальным расположением жгутиков в таком положении находится, смотря по обстоятельствам, то один, то другой пучок.
Перитрихально расположенные жгутики Escherichia coli
работают как один хорошо скоординированный спиральный пучок и проталкивают клетку через среду. В тех случаях, когда направление вращения отдельных жгутиков меняется, клетка начинает «кувыркаться». По-видимому, перитрихально расположенные жгутики не могут служить тянущим приспособлением.
Бактерии, снабженные жгутиками, могут передвигаться очень быстро: Bacillus megaterium
со скоростью 1,6 мм/мин, Vibrio cholerae -
12 мм/мин. Это соответствует примерно от 300 до 3000 длин тела в минуту.
Тонкое строение жгутиков.Жгутики представляют собой спирально закрученные нити. У разных бактерий они различаются по своей толщине (12-18 нм), длине (до 20 мкм), а также по длине и амплитуде витка. Эти параметры характерны для каждого вида. У некоторых бактерий могут образовываться жгутики разных типов. Нити жгутиков состоят из специфического белка флагеллина. Они построены из субъединиц с относительно малой молекулярной массой. Субьединицы располагаются по спирали вокруг внутреннего свободного пространства (подобно белковым молекулам в вирусе табачной мозаики). Таким образом, структура жгутика определяется свойствами белковых субъединиц.
Жгутик состоит из трех частей - описанной выше спиральной нити, «крюка» вблизи поверхности клетки и базального тельца. С помощью базального тельца жгутик закреплен в плазматической мембране и в клеточной стенке (рис. 2.38). Оно состоит из центрального стержня, на котором у грам-отрицательных бактерий находятся две пары колец. Наружная пара (кольца L и Р) расположены на уровне наружного и внутреннего слоев клеточной стенки, а внутренняя пара (кольца S и М) - на уровне наружного слоя плазматической мембраны. Так как у грам-положительных бактерий наружная пара колец отсутствует, полагают, что для вращения жгутиков необходима только внутренняя пара. Можно представить себе, что кольцо М действует как приводной диск, а кольцо S играет роль подшипника на внутренней поверхности пептидогликанового слоя. Молекулярный механизм вращательного «мотора» жгутика пока не выяснен.
 О- и Н-аитигены.Proteus vulgaris
часто распространяется по всей поверхности агара в виде тонкого серого налета (Н-форма, от нем. Hauch - налет). Такое «роение» объясняется большой подвижностью клеток. Некоторые штаммы налета не образуют (О-форма, от нем. ohne Hauch - без налета). Эти штаммы неподвижны, они лишены жгутиков. Отсюда ведет свое начало обычная терминология, принятая в бактериальной серодиагностике; антигены поверхности или вообще тела клетки (соматические) называют О-антигенами, а антигены жгутиков - Н-антигенами.
Фимбрии и пили.Поверхность некоторых бактерий покрыта большим числом (от 10 до нескольких тысяч) длинных, тонких прямых нитей толщиной 3-25 нм и длиной до 12 мкм, называемых фимбриями или пилями. Они встречаются как у жгутиконосных видов, так и у форм, лишенных жгутиков. От них следует отличать половые пили, или пили типа F, которые были обнаружены у клеток - доноров Escherichia coli
К 12, т.е. у штаммов, содержащих половой фактор F (F + , Hfr). Пили F встречаются только по одной или по две на клетку, они имеют вид полых белковых трубочек длиной от 0,5 до 10 мкм.
Хемотаксис.Свободно передвигающиеся бактерии способны к таксисам - направленным движениям, определяемым внешними стимулами. В зависимости от факторов среды, вызывающих направленное движение, говорят о хемотаксисе, аэротаксисе, фототаксисе и магнитотаксисе.
Подвижные бактерии реагируют на химические раздражители - скапливаются в одних местах, а других мест избегают. Такая реакция свободно передвигающихся организмов называется хемотаксисом. Скопления бактерий образуются под действием химических факторов следующим образом (рис. 2.39). У форм с перитрихальными жгутиками возможны только два типа двигательного поведения: прямолинейное движение и кувыркание. Последнее прерывает прямолинейную пробежку и изменяет направление пути. Когда бактерия оказывается в среде с градиентом концентрации «привлекающего» ее субстрата (аттрактанта), ее прямолинейное движение длится многие секунды, если она плывет по направлению к оптимальной его концентрации; однако такое движение через несколько секунд прекратится, если бактерия плывет в противоположном направлении. Хотя направление прямолинейного движения после кувыркания оказывается совершенно случайным, тем не менее зависимость длительности такого движения от его направления приводит в конечном результате к накоплению бактерий в области оптимальной концентрации субстрата. За чувствительность к химическому стимулу и за реагирование на него ответственны хеморецепторы. В ряде случаев эти хеморецепторы действуют независимо от способности бактерий утилизировать данный субстрат. Например, некоторые мутанты продолжают совершенно нормально реагировать на определенное питательное вещество, хотя и потеряли способность его использовать.
О- и Н-аитигены.Proteus vulgaris
часто распространяется по всей поверхности агара в виде тонкого серого налета (Н-форма, от нем. Hauch - налет). Такое «роение» объясняется большой подвижностью клеток. Некоторые штаммы налета не образуют (О-форма, от нем. ohne Hauch - без налета). Эти штаммы неподвижны, они лишены жгутиков. Отсюда ведет свое начало обычная терминология, принятая в бактериальной серодиагностике; антигены поверхности или вообще тела клетки (соматические) называют О-антигенами, а антигены жгутиков - Н-антигенами.
Фимбрии и пили.Поверхность некоторых бактерий покрыта большим числом (от 10 до нескольких тысяч) длинных, тонких прямых нитей толщиной 3-25 нм и длиной до 12 мкм, называемых фимбриями или пилями. Они встречаются как у жгутиконосных видов, так и у форм, лишенных жгутиков. От них следует отличать половые пили, или пили типа F, которые были обнаружены у клеток - доноров Escherichia coli
К 12, т.е. у штаммов, содержащих половой фактор F (F + , Hfr). Пили F встречаются только по одной или по две на клетку, они имеют вид полых белковых трубочек длиной от 0,5 до 10 мкм.
Хемотаксис.Свободно передвигающиеся бактерии способны к таксисам - направленным движениям, определяемым внешними стимулами. В зависимости от факторов среды, вызывающих направленное движение, говорят о хемотаксисе, аэротаксисе, фототаксисе и магнитотаксисе.
Подвижные бактерии реагируют на химические раздражители - скапливаются в одних местах, а других мест избегают. Такая реакция свободно передвигающихся организмов называется хемотаксисом. Скопления бактерий образуются под действием химических факторов следующим образом (рис. 2.39). У форм с перитрихальными жгутиками возможны только два типа двигательного поведения: прямолинейное движение и кувыркание. Последнее прерывает прямолинейную пробежку и изменяет направление пути. Когда бактерия оказывается в среде с градиентом концентрации «привлекающего» ее субстрата (аттрактанта), ее прямолинейное движение длится многие секунды, если она плывет по направлению к оптимальной его концентрации; однако такое движение через несколько секунд прекратится, если бактерия плывет в противоположном направлении. Хотя направление прямолинейного движения после кувыркания оказывается совершенно случайным, тем не менее зависимость длительности такого движения от его направления приводит в конечном результате к накоплению бактерий в области оптимальной концентрации субстрата. За чувствительность к химическому стимулу и за реагирование на него ответственны хеморецепторы. В ряде случаев эти хеморецепторы действуют независимо от способности бактерий утилизировать данный субстрат. Например, некоторые мутанты продолжают совершенно нормально реагировать на определенное питательное вещество, хотя и потеряли способность его использовать.
 Аэротаксис.У подвижных бактерий можно определить тип метаболизма (аэробный или анаэробный) по их аэротаксическим движениям и скоплению на определенных расстояниях от края покровного стекла. В слое бактерий, помещенных между предметным и покровным стеклами, аэрофильные бактерии скапливаются у края покровного стекла или в непосредственной близости от оказавшихся в препарате пузырьков воздуха; это указывает на их потребность в аэробных условиях и на то, что необходимую энергию они получают за счет дыхания (рис. 2.40). Строго анаэробные бактерии будут скапливаться в центре. Микроаэрофильные бактерии, например некоторые псевдомонады и спириллы, будут держаться на определенном расстоянии от края. С помощью бактерий, проявляющих положительный аэротаксис, Энгельману удалось продемонстрировать выделение кислорода локально освещаемыми хлоропластами зеленой водоросли Spirogyra.
Аэротаксис.У подвижных бактерий можно определить тип метаболизма (аэробный или анаэробный) по их аэротаксическим движениям и скоплению на определенных расстояниях от края покровного стекла. В слое бактерий, помещенных между предметным и покровным стеклами, аэрофильные бактерии скапливаются у края покровного стекла или в непосредственной близости от оказавшихся в препарате пузырьков воздуха; это указывает на их потребность в аэробных условиях и на то, что необходимую энергию они получают за счет дыхания (рис. 2.40). Строго анаэробные бактерии будут скапливаться в центре. Микроаэрофильные бактерии, например некоторые псевдомонады и спириллы, будут держаться на определенном расстоянии от края. С помощью бактерий, проявляющих положительный аэротаксис, Энгельману удалось продемонстрировать выделение кислорода локально освещаемыми хлоропластами зеленой водоросли Spirogyra.
 Фототаксис. Фототрофным пурпурным бактериям для получения энергии необходим свет. Не удивительно поэтому, что в результате фототаксиса они скапливаются в освещенном месте. Если выдержать в темноте препарат, в котором плотная суспензия клеток Chromatium будет равномерно распределена под покровным стеклом, а затем направить на него сфокусированный пучок света, то бактерии сосредоточатся в области светового пятна. Клетки, попавшие в это пятно случайно в результате своего беспорядочного движения, уже не могут его покинуть. Как только они попадут в темную зону, направление движения жгутиков мгновенно меняется на обратное и клетки возвращаются в освещенное место. Изменение работы жгутиков происходит так быстро, что эта реакция получила название «реакция испуга» (фоботаксис). Впрочем, для того чтобы вызвать такой ответ, достаточно даже небольшого различия в освещенности двух участков. Мелкие клетки Chromatium скапливаются уже в таком месте, где освещенность всего на 0,7% выше, чем в окружающей области. Таким образом, по своей чувствительности к световому контрасту они приближаются к сетчатке человеческого глаза (для которой соответствующий порог равен 0,4%).
Магиитотаксис. Из поверхностных слоев донного ила пресноводных водоемов, а также морей были выделены бактерии (палочки, спириллы, кокки), способные ориентироваться в магнитном поле и перемещаться в направлении линий магнитного поля. Они содержат много железа (0,4% сухого вещества) в форме ферромагнитной окиси железа (магнетита), которая находится в гранулах (магнитосомах), расположенных около мест прикрепления жгутиков. Бактерии, выделенные в северном полушарии, «ищут» север; здесь линии магнитного поля проходят под углом около 70° к горизонту вниз, вглубь водоема. Магнитотаксическоё поведение направляет бактерии в глубину ила, где очень мало или вовсе нет кислорода. Так как магнитотаксические бактерии - анаэробы или микроаэрофилы, их реакция на магнитное поле понятна с точки зрения экологии. Такие клетки, завезенные в южное полушарие, в массе своей, конечно, погибнут; выживут лишь немногие «неправильно» поляризованные клетки, которые могут затем размножиться. Полярность, очевидно, генетически не зафиксирована.
Фототаксис. Фототрофным пурпурным бактериям для получения энергии необходим свет. Не удивительно поэтому, что в результате фототаксиса они скапливаются в освещенном месте. Если выдержать в темноте препарат, в котором плотная суспензия клеток Chromatium будет равномерно распределена под покровным стеклом, а затем направить на него сфокусированный пучок света, то бактерии сосредоточатся в области светового пятна. Клетки, попавшие в это пятно случайно в результате своего беспорядочного движения, уже не могут его покинуть. Как только они попадут в темную зону, направление движения жгутиков мгновенно меняется на обратное и клетки возвращаются в освещенное место. Изменение работы жгутиков происходит так быстро, что эта реакция получила название «реакция испуга» (фоботаксис). Впрочем, для того чтобы вызвать такой ответ, достаточно даже небольшого различия в освещенности двух участков. Мелкие клетки Chromatium скапливаются уже в таком месте, где освещенность всего на 0,7% выше, чем в окружающей области. Таким образом, по своей чувствительности к световому контрасту они приближаются к сетчатке человеческого глаза (для которой соответствующий порог равен 0,4%).
Магиитотаксис. Из поверхностных слоев донного ила пресноводных водоемов, а также морей были выделены бактерии (палочки, спириллы, кокки), способные ориентироваться в магнитном поле и перемещаться в направлении линий магнитного поля. Они содержат много железа (0,4% сухого вещества) в форме ферромагнитной окиси железа (магнетита), которая находится в гранулах (магнитосомах), расположенных около мест прикрепления жгутиков. Бактерии, выделенные в северном полушарии, «ищут» север; здесь линии магнитного поля проходят под углом около 70° к горизонту вниз, вглубь водоема. Магнитотаксическоё поведение направляет бактерии в глубину ила, где очень мало или вовсе нет кислорода. Так как магнитотаксические бактерии - анаэробы или микроаэрофилы, их реакция на магнитное поле понятна с точки зрения экологии. Такие клетки, завезенные в южное полушарие, в массе своей, конечно, погибнут; выживут лишь немногие «неправильно» поляризованные клетки, которые могут затем размножиться. Полярность, очевидно, генетически не зафиксирована.
6. Жгутики, ворсинки, пили. Способы передвижения прокариот. Методы определения подвижности у бактерий
Жгутики – тонкие, длинные, нитевидные, белковые образования. Из белка лабеллина. Он обладает сократительной способностью. По хар-ру расположен жгутиков и их кол-ву различ: монотрихи (один полярно расположенный жгутик), лофотрихи (пучёк жгутиков на одном конце), анфитрихи (по 1 или пучёк на противоположных концах кл), перитрихи (по всей поверхности), атрихи (неподвижные). Они хар-ны для молодых культур, с возрастом или при недостатке пит ср жгутики утрачиваются. Подвижность бактер определ микро и макроскопич методами. При микроскопич готовят мазки раздавленной или висячей капли. макроскопич – методом укола, посевом на полужиткий агар. Жгутики сост из 3 компонентов: базальное тельце, крючок, спиральная жгутиковая нить. Баз тельце сост из системы особых колец. У гр- бактер их 2 пары: внешняя L и Р и внутрен S и М. У гр+ S и М. в рез-те их вращения относительно друг друга происходит вращение жгутика. Крючок – изогнутый белковый цилиндр, выполняющий ф-цию гибкого связывающего звена между базальным тельцем и жёсткой нитью жгутика. Ьазальное тельце – сложная структура, состоящая из центрального стержня и колец. Передвижение прокариот осуществляется вращательными, поступательными, сгибательными движениями. Пили. У бактер, являющихся носителями плазмид имеются нитевидные структуры белковой природы. Построены из белка пиллина. Синтез этих ворсинок находится под контролем плазмидных генов. Пили явл аппаратом конъюгации с их помощью устанавливается контакт между кл донорам и кл реципиентом. Существует 2 класса пилей – половые и общего типа (фимбрии). Секс-пили – 1,2 и более 5 на 1 кл. Ворсинки (фимбрии). Короткие нити, кол-во кот может достигать много тыс. с их помощью бактерии прикрепляются к определённым поверхностям. Для многих болезнетворных бактерий фибрии явл фактором патогенности, т.к. с их помощью бактерии прикрепляются к чуствит кл и явл ф-ром адгизии. Вызывают агглютинацию эритроцитов.
7.Простые и сложные методы окрашивания микроорган
Практическое значение. Препараты окрашивают простым и сложным методами. Простой метод. Для окраски используют какой-либо один красящий р-р. На фиксированный мазок наносят р-р одного красителя: метиленовым синим окрашив 4 мин, генцианвиолетом – 2 мин, фуксином – 1 мин. Краску смывают водой, мазок высушивают фильтровальной бумагой. На готовый мазок наносят каплю иммерсионного масла, микроскопируют. Простая окраска позволяет быстро ознакомится с морфологией бактерий. Сложные методы. Применяют несколько р-ров красителе и реактивов. Они позволяют определить морфологию бактерий, их тинкториальные особенности и наличие структурных элементов кл, что имеет важное дифференциально-диагностическое значение. Одним из методов явл окрашивание по Грамму: на фиксированный препарат на 2 мин накладывают фильтровальную бумагу, пропитанную генцианвиолетом. Бумажку снимают и наносят р-р Люголя на 2 мин. Сливают, мазок обрабатывают спиртом 30 сек, промывают водой и окрашивают фуксином 1 мин. По рез-ту окрашивания определ тип клеточной стенки. Методы окраски спор: метод Меллера: фиксированный мазок протравляют хромовой к-той 2 мин, промывают водой, высушивают фильтровальной бумагой, накладывают фильтровальн бумагу на мазок и наносят фуксин, препарат подогревают, окрашивают 7 мин, бумажку и краску сливают и обрабатывают серной к-той 5 сек, промывают водой, окрашив метиленовой синью 4 мин, промывают водой, просушивают фильтровальной бумагой. Микроскопируют: споры розово-красные, вегетативная кл – синяя. Метод Златогорова – такой же, только не обрабатыв хромовой к-той. Метод Пешкова: мазок фиксируют, окрашивают метиленовой синью с подогревом, смывают водой, докрашивают р-ром нейтрального Красного 10 сек, смывают водой, высушивают фильтровальной бумагой. Споры синие, кл – красная.
8. Культивирование аэробных микроорган в условиях лаборатории. Методы выделения чистой культуры аэробных микроорган
Микроорганизмы, выращенные на искусственных пит ср – микробными культурами, а получение их роста на пит ср - культивированием. Для культивирования необходимы условия: оптимальный температурный режим с учетом, к какой группе относится исследуемый вид бактерий, соответствующие пит ср, аэробиоз (или анаэробиоз). Для обеспечения постоянной оптимальной температуры служат термостаты. Лабораторный термостат - шкаф с двойными стенками, снаружи облицованный материалом непроводником тепла (пластик), внутренняя стенка металлическая. Между двумя металлическими стенками находится вода (или воздух), подогреваемая электричеством. От нагретой воды через внутреннюю металлическую стенку тепло поступает в термостат. Внутри имеются сетчатые полочки, на которых размещают штативы с пробирками, чашки Петри и др. Постоянная температура поддерживается при помощи терморегуляторов - при достижении температуры заданного уровня автоматически происходит отключение прибора; при снижении температуры термостат вновь включается автоматически. Помимо обеспечения температурного режима, следует учитывать тип дыхания микроорганизмов: при аэробном типе дыхания никаких дополнительных условий создавать не нужно. Выделение из смеси одного вида микроба – выделение чистой культуры. Один из первых методов предложил Пастер – метод разведения. Исследуемый материал последовательно разводят в жидкой пит среде: берут ряд пробирок с МПБ, исследуемый материал вносят в первую пробирку, перемешивают, из неё переносят во вторую и т.д. Пастер предполагал, что в последней пробирке возможен рост одного вида микроба. Но это не так. Метод Коха – применяется плотная среда – используя принцип Пастера, исследуемый материал разводят в 4-5 пробирках с расплавленным и остужённым МПА, осторожно содержимое пробирки выливают в чашку Петри и распределяют среду тонким слоем, чашку закрывают, и когда А остынет переворачивают вверх дном. Ставят в термостат. Там где концентрация микробов меньше вырастают изолированные друг от друга колонии. С обратной стороны отмечают нужную колонию, делают посевы на МПБ и МПА и вырастает чистая культура. Метод Дригальского – метод пластинчатого посева. берут 4-5 чашек Петри. Агаровую среду расплавляют в колбе, разливают в чашки и ставят в термостат вверх дном. Шпателем Дригальского или пастеровской пипеткой равномерно растирают на поверхности среды каплю. Этим же шпателем растирают на поверхности второй чашки и т.д. Помещают в термостат вверх дном. Нужную культуру засевают в МПА и МПБ. Биологический метод – исследуемый материал вводят восприимчивому жив. При наличии патогенного микроба жив гибнут, их вскрывают и делают посевы. Метод Шукевича – подвижный микроб переходит на поверхность А из конденсационной жидкости, из верхнего края выросшей культуры делают посевы и получают чистую культуру. Химический метод – к пит ср добавляют хим в-ва, кот действуют на одних убийственно, у других задерживается рост, а третьи не восприимчивы.
9.Химический состав бактериальной Кл
75-85% вода, 25-15% - сухой остаток. Ведущая роль принадлежит 4 осн элемента, кот получил наз органогены: кислород – 30% сухого остатка, Н – 6-8%, С-45-55%, N-8-15%. Вода в кл может быть в 2 состояниях: свободная вода, кот явл растворит для кристалич в-в и в ней происходит движение ионов; связанная вода, кот входит в состав белков, жиров и углеводов. В бактериологич кл на долю белков – 60-70%, углеводы – 20%, липиды -1-2%. Повышенное содержан липидов придаёт клетке кислотно-спирто-щелочеустойчивость. Белки – высокомолекулярные полимерные соединения, образующиеся при гидролизе аминок-ты – сложные – протеиды, простые - протеины. Ф-ции белков – гл структурный материал для всех клеточн мембран, они обеспечив двигательн ф-ции, транспорт питат в-в через мембрану. Углеводы – многоатомные спирты (манит, дульцит) и полисахариды (гликоген). Играют энергитич роль в кл. Липиды – жирные к-ты и нейтральные жиры, фосфолипиды цитоплазматич мембраны. Явл резервом кл, использ как исходн компонент для синтеза белка. Мин в-ва – 3-10% сух остатка. Микро и макроэлементы. Микробные ф-ты. Гл св-ва: специфичность и термолябильность. У микроорганизм набор ф-тов генетически закреплён и передаётся по наследству. Различ ф-ты: 1. экзоф-ты – выдел кл во внешн среду и катализир разложен сложных в-в субстрата до более простых. 2. эндоф-ты – локализуются в самой кл и участвует во внутриклеточн процессах обмена в-в. 3. конститутивные – явл постоян компонентом кл и могут быть обнаружены даже при отсутствии в среде субстрата, кот они катализируют. 4. адаптивные – вырабатываются кл только тогда, когда в среде появл соотвествующий субстрат. Различают: оксиредуктазу, трансферазу, гидролазу, лиазу, изомеразу, лигазу и киназу. Наличие ф-тов можно обнаружить с помощью спец сред.




Два других отдела, отделенных мембраной, пирреллюлозому или рибоплазму, которая содержит рибосомы и связанные белки, и свободный от рибосом парафоплазму (Glockner, 2003). 3. Характеристика общих свойств микроорганизмов Микроорганизмы - это организмы, невидимые невооруженным глазом из-за их незначительных размеров. Этот критерий - единственный, который их объединяет. В остальном мир...

По числу совпадающих признаков. Указанный подход к систематике микроорганизмов достаточно объективен, однако для его реализации необходимы обширные математические расчеты с использованием электронно-вычислительных машин. После подробного изучения микроорганизму дают научное название, которое должно быть выражено двумя латинскими словами, как этого требует биноминальная номенклатура, предложенная...
В автотрофных условиях. Соответственно окисляемым субстратам выделяют такие группы хемолитоавтотрофов, как водородные, нитрифицирующие, серные бактерии, железобактерии. К числу хемолитоавтотрофных микроорганизмов, окисляющих Н2, относятся также многие метанобразующие бактерии, отдельные представители ацетатобразующих, сульфат- и серувосстанавливающих бактерий. Различные возможности проявляют...





Высказывается также предположение, что мезосомы не принимают активного участия в процессах клеточного метаболизма, но выполняют структурную функцию, обеспечивая компартментализацию прокариотной клетки, т. е. пространственное разграничение внутриклеточного содержимого на относительно обособленные отсеки, что создает более благоприятные условия для протекания определенных последовательностей...
Бактерии — самая древняя группа организмов из ныне существующих на Земле. Первые бактерии появились, вероятно, более 3,5 млрд лет назад и на протяжении почти миллиарда лет были единственными живыми существами на нашей планете. Поскольку это были первые представители живой природы, их тело имело примитивное строение.
Со временем их строение усложнилось, но и поныне бактерии считаются наиболее примитивными одноклеточными организмами. Интересно, что некоторые бактерии и сейчас ещё сохранили примитивные черты своих древних предков. Это наблюдается у бактерий, обитающих в горячих серных источниках и бескислородных илах на дне водоёмов.
Большинство бактерий бесцветно. Только немногие окрашены в пурпурный или в зелёный цвет. Но колонии многих бактерий имеют яркую окраску, которая обусловливается выделением окрашенного вещества в окружающую среду или пигментированием клеток.
Первооткрывателем мира бактерий был Антоний Левенгук — голландский естествоиспытатель 17 века, впервые создавший совершенную лупу-микроскоп, увеличивающую предметы в 160-270 раз.
Бактерии относят к прокариотам и выделяют в отдельное царство — Бактерии.
Форма тела
Бактерии — многочисленные и разнообразные организмы. Они различаются по форме.
| Название бактерии | Форма бактерии | Изображение бактерии |
| Кокки | Шарообразная | |
| Бацилла |  | Палочковидная |
| Вибрион | Изогнутая в виде запятой | |
| Спирилла |  | Спиралевидная |
| Стрептококки | 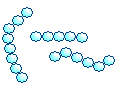 | Цепочка из кокков |
| Стафилококки |  | Грозди кокков |
| Диплококки | Две круглые бактерии, заключённые в одной слизистой капсуле |
Способы передвижения
Среди бактерий есть подвижные и неподвижные формы. Подвижные передвигаются за счёт волнообразных сокращений или при помощи жгутиков (скрученные винтообразные нити), которые состоят из особого белка флагеллина. Жгутиков может быть один или несколько. Располагаются они у одних бактерий на одном конце клетки, у других — на двух или по всей поверхности.

Но движение присуще и многим иным бактериям, у которых жгутики отсутствуют. Так, бактерии, покрытые снаружи слизью, способны к скользящему движению.
У некоторых лишённых жгутиков водных и почвенных бактерий в цитоплазме имеются газовые вакуоли. В клетке может быть 40-60 вакуолей. Каждая из них заполнена газом (предположительно — азотом). Регулируя количество газа в вакуолях, водные бактерии могут погружаться в толщу воды или подниматься на её поверхность, а почвенные бактерии — передвигаться в капиллярах почвы.
Место обитания
В силу простоты организации и неприхотливости бактерии широко распространены в природе. Бактерии обнаружены везде: в капле даже самой чистой родниковой воды, в крупинках почвы, в воздухе, на скалах, в полярных снегах, песках пустынь, на дне океана, в добытой с огромной глубины нефти и даже в воде горячих источников с температурой около 80ºС. Обитают они на растениях, плодах, у различных животных и у человека в кишечнике, ротовой полости, на конечностях, на поверхности тела.
Бактерии — самые мелкие и самые многочисленные живые существа. Благодаря малым размерам они легко проникают в любые трещины, щели, поры. Очень выносливы и приспособлены к различным условиям существования. Переносят высушивание, сильные холода, нагревание до 90ºС, не теряя при этом жизнеспособность.
Практически нет места на Земле, где не встречались бы бактерии, но в разных количествах. Условия жизни бактерий разнообразны. Одним из них необходим кислород воздуха, другие в нём не нуждаются и способны жить в бескислородной среде.
В воздухе: бактерии поднимаются в верхние слои атмосферы до 30 км. и больше.
Особенно много их в почве. В 1 г. почвы могут содержаться сотни миллионов бактерий.
В воде: в поверхностных слоях воды открытых водоёмов. Полезные водные бактерии минерализуют органические остатки.
В живых организмах: болезнетворные бактерии попадают в организм из внешней среды, но лишь в благоприятных условиях вызываю заболевания. Симбиотические живут в органах пищеварения, помогая расщеплять и усваивать пищу, синтезируют витамины.
Внешнее строение
Клетка бактерии одета особой плотной оболочкой — клеточной стенкой, которая выполняет защитную и опорную функции, а также придаёт бактерии постоянную, характерную для неё форму. Клеточная стенка бактерии напоминает оболочку растительной клетки. Она проницаема: через неё питательные вещества свободно проходят в клетку, а продукты обмена веществ выходят в окружающую среду. Часто поверх клеточной стенки у бактерий вырабатывается дополнительный защитный слой слизи — капсула. Толщина капсулы может во много раз превышать диаметр самой клетки, но может быть и очень небольшой. Капсула — не обязательная часть клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она предохраняет бактерию от высыхания.
На поверхности некоторых бактерий имеются длинные жгутики (один, два или много) или короткие тонкие ворсинки. Длина жгутиков может во много раз превышать разметы тела бактерии. С помощью жгутиков и ворсинок бактерии передвигаются.

Внутреннее строение
Внутри клетки бактерии находится густая неподвижная цитоплазма. Она имеет слоистое строение, вакуолей нет, поэтому различные белки (ферменты) и запасные питательные вещества размещаются в самом веществе цитоплазмы. Клетки бактерий не имеют ядра. В центральной части их клетки сконцентрировано вещество, несущее наследственную информации. Бактерии, — нуклеиновая кислота — ДНК. Но это вещество не оформлено в ядро.
Внутренняя организация бактериальной клетки сложна и имеет свои специфические особенности. Цитоплазма отделяется от клеточной стенки цитоплазматической мембраной. В цитоплазме различают основное вещество, или матрикс, рибосомы и небольшое количество мембранных структур, выполняющих самые различные функции (аналоги митохондрий, эндоплазматической сети, аппарата Гольджи). В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Гранулы могут состоять из соединений, которые служат источником энергии и углерода. В бактериальной клетке встречаются и капельки жира.
В центральной части клетки локализовано ядерное вещество — ДНК, не отграниченная от цитоплазмы мембраной. Это аналог ядра — нуклеоид. Нуклеоид не обладает мембраной, ядрышком и набором хромосом.
Способы питания
У бактерий наблюдаются разные способы питания. Среди них есть автотрофы и гетеротрофы. Автотрофы — организмы, способные самостоятельно образовывать органические вещества для своего питания.
Растения нуждаются в азоте, но сами усваивают азот воздуха не могут. Некоторые бактерии соединяют содержащиеся в воздухе молекулы азота с другими молекулами, в результате чего получаются вещества, доступные для растений.
Эти бактерии поселяются в клетках молодых корней, что приводит к образованию на корнях утолщений, называемых клубеньками. Такие клубеньки образуются на корнях растений семейства бобовых и некоторых других растений.

Корни дают бактериям углеводы, а бактерии корням — такие содержащие азот вещества, которые могут быть усвоены растением. Их сожительство взаимовыгодно.
Корни растений выделяют много органических веществ (сахара, аминокислоты и другие), которыми питаются бактерии. Поэтому в слое почвы, окружающем корни, поселяется особенно много бактерий. Эти бактерии превращают отмершие остатки растений в доступные для растения вещества. Этот слой почвы называют ризосферой.
Существует несколько гипотез о проникновении клубеньковых бактерий в ткани корня:
- через повреждения эпидермальной и коровой ткани;
- через корневые волоски;
- только через молодую клеточную оболочку;
- благодаря бактериям-спутникам, продуцирующим пектинолитические ферменты;
- благодаря стимуляции синтеза В-индолилуксусной кислоты из триптофана, всегда имеющегося в корневых выделениях растений.
Процесс внедрения клубеньковых бактерий в ткань корня состоит из двух фаз:
- инфицирование корневых волосков;
- процесс образования клубеньков.
В большинстве случаев внедрившаяся клетка, активно размножается, образует так называемые инфекционные нити и уже в виде таких нитей перемещается в ткани растения. Клубеньковые бактерии, вышедшие из инфекционной нити, продолжают размножаться в ткани хозяина.

Наполняющиеся быстро размножающимися клетками клубеньковых бактерий растительные клетки начинают усиленно делиться. Связь молодого клубенька с корнем бобового растения осуществляется благодаря сосудисто-волокнистым пучкам. В период функционирования клубеньки обычно плотные. К моменту проявления оптимальной активности клубеньки приобретают розовую окраску (благодаря пигменту легоглобину). Фиксировать азот способны лишь те бактерии, которые содержат легоглобин.
Бактерии клубеньков создают десятки и сотни килограммов азотных удобрений на гектаре почвы.
Обмен веществ
Бактерии отличаются друг от друга обменом веществ. У одних он идёт при участии кислорода, у других — без его участия.
Большинство бактерий питается готовыми органическими веществами. Лишь некоторые из них (сине-зелёные, или цианобактерии), способны создавать органические вещества из неорганических. Они сыграли важную роль в накоплении кислорода в атмосфере Земли.
Бактерии впитывают вещества извне, разрывают их молекулы на части, из этих частей собирают свою оболочку и пополняют своё содержимое (так они растут), а ненужные молекулы выбрасывают наружу. Оболочка и мембрана бактерии позволяет ей впитывать только нужные вещества.

Если бы оболочка и мембрана бактерии были полностью непроницаемыми, в клетку не попали бы никакие вещества. Если бы они были проницаемыми для всех веществ, содержимое клетки перемешалось бы со средой — раствором, в которой обитает бактерия. Для выживания бактерии необходима оболочка, которая нужные вещества пропускает, а ненужные — нет.
Бактерия поглощает находящиеся близ неё питательные вещества. Что происходит потом? Если она может самостоятельно передвигаться (двигая жгутик или выталкивая назад слизь), то она перемещается, пока не найдёт необходимые вещества.
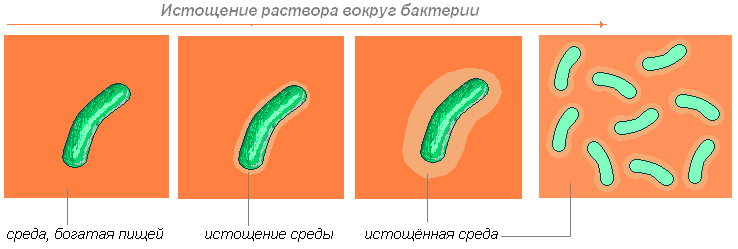
Если она двигаться не может, то ждёт, пока диффузия (способность молекул одного вещества проникать в гущу молекул другого вещества) не принесёт к ней необходимые молекулы.
Бактерии в совокупности с другими группами микроорганизмов выполняют огромную химическую работу. Превращая различные соединения, они получают необходимую для их жизнедеятельности энергию и питательные вещества. Процессы обмена веществ, способы добывания энергии и потребности в материалах для построения веществ своего тела у бактерий разнообразны.
Другие бактерии все потребности в углероде, необходимом для синтеза органических веществ тела, удовлетворяют за счёт неорганических соединений. Они называются автотрофами. Автотрофные бактерии способны синтезировать органические вещества из неорганических. Среди них различают:
Хемосинтез
Использование лучистой энергии — важнейший, но не единственный путь создания органического вещества из углекислого газа и воды. Известны бактерии, которые в качестве источника энергии для такого синтеза используют не солнечный свет, а энергию химических связей, происходящих в клетках организмов при окислении некоторых неорганических соединений — сероводорода, серы, аммиака, водорода, азотной кислоты, закисных соединений железа и марганца. Образованное с использованием этой химической энергии органическое вещество они используют для построения клеток своего тела. Поэтому такой процесс называют хемосинтезом.
Важнейшую группу хемосинтезирующих микроорганизмов составляют нитрифицирующие бактерии. Эти бактерии живут в почве и осуществляют окисление аммиака, образовавшегося при гниении органических остатков, до азотной кислоты. Последняя, реагирует с минеральными соединениями почвы, превращаются в соли азотной кислоты. Этот процесс проходит в две фазы.
Железобактерии превращают закисное железо в окисное. Образованная гидроокись железа оседает и образует так называемую болотную железную руду.
Некоторые микроорганизмы существуют за счёт окисления молекулярного водорода, обеспечивая тем самым автотрофный способ питания.
Характерной особенностью водородных бактерий является способность переключаться на гетеротрофный образ жизни при обеспечении их органическими соединениями и отсутствии водорода.
Таким образом, хемоавтотрофы являются типичными автотрофами, так как самостоятельно синтезируют из неорганических веществ необходимые органические соединения, а не берут их в готовом виде от других организмов, как гетеротрофы. От фототрофных растений хемоавтотрофные бактерии отличаются полной независимостью от света как источника энергии.
Бактериальный фотосинтез
Некоторые пигментосодержащие серобактерии (пурпурные, зелёные), содержащие специфические пигменты — бактериохлорофиллы, способны поглощать солнечную энергию, с помощью которой сероводород в их организмах расщепляется и отдаёт атомы водорода для восстановления соответствующих соединений. Этот процесс имеет много общего с фотосинтезом и отличается только тем, что у пурпурных и зелёных бактерий донором водорода является сероводород (изредка — карбоновые кислоты), а у зелёных растений — вода. У тех и других отщепление и перенесение водорода осуществляется благодаря энергии поглощённых солнечных лучей.
Такой бактериальный фотосинтез, который происходит без выделения кислорода, называется фоторедукцией. Фоторедукция углекислого газа связана с перенесением водорода не от воды, а от сероводорода:
6СО 2 +12Н 2 S+hv → С6Н 12 О 6 +12S=6Н 2 О
Биологическое значение хемосинтеза и бактериального фотосинтеза в масштабах планеты относительно невелико. Только хемосинтезирующие бактерии играют существенную роль в процессе круговорота серы в природе. Поглощаясь зелёными растениями в форме солей серной кислоты, сера восстанавливается и входит в состав белковых молекул. Далее при разрушении отмерших растительных и животных остатков гнилостными бактериями сера выделяется в виде сероводорода, который окисляется серобактериями до свободной серы (или серной кислоты), образующий в почве доступные для растения сульфиты. Хемо- и фотоавтотрофные бактерии имеют существенное значение в круговороте азота и серы.
Спорообразование
Внутри бактериальной клетки образуются споры. В процессе спорообразования бактериальная клетка претерпевает ряд биохимических процессов. В ней уменьшается количество свободной воды, снижается ферментативная активность. Это обеспечивает устойчивость спор к неблагоприятным условиям внешней среды (высокой температуре, высокой концентрации солей, высушиванию и др.). Спорообразование свойственно только небольшой группе бактерий.

Споры — не обязательная стадия жизненного цикла бактерий. Спорообразование начинается лишь при недостатке питательных веществ или накоплении продуктов обмена. Бактерии в виде спор могут длительное время находиться в состоянии покоя. Споры бактерий выдерживают продолжительное кипячение и очень длительное проммораживание. При наступлении благоприятных условий спора прорастает и становится жизнеспособной. Спора бактерий — это приспособление к выживанию в неблагоприятных условиях.
Размножение
Размножаются бактерии делением одной клетки на две. Достигнув определённого размера, бактерия делится на две одинаковые бактерии. Затем каждая из них начинает питаться, растёт, делится и так далее.

После удлинения клетки постепенно образуется поперечная перегородка, а затем дочерние клетки расходятся; у многих бактерий в определённых условиях клетки после деления остаются связанными в характерные группы. При этом в зависимости от направления плоскости деления и числа делений возникают разные формы. Размножение почкованием встречается у бактерий как исключение.
При благоприятных условиях деление клеток у многих бактерий происходит через каждые 20-30 минут. При таком быстром размножении потомство одной бактерии за 5 суток способно образовать массу, которой можно заполнить все моря и океаны. Простой подсчёт показывает, что за сутки может образоваться 72 поколения (720 000 000 000 000 000 000 клеток). Если перевести в вес — 4720 тонн. Однако в природе этого не происходит, так как большинство бактерий быстро погибают под действием солнечного света, при высушивании, недостатке пищи, нагревании до 65-100ºС, в результате борьбы между видами и т.д.
Бактерия (1), поглотившая достаточно пищи, увеличивается в размерах (2) и начинает готовиться к размножению (делению клетки). Её ДНК (у бактерии молекула ДНК замкнута в кольцо) удваивается (бактерия производит копию этой молекулы). Обе молекулы ДНК (3,4) оказываются, прикреплены к стенке бактерии и при удлинении бактерии расходятся в стороны (5,6). Сначала делится нуклеотид, затем цитоплазма.

После расхождения двух молекул ДНК на бактерии появляется перетяжка, которая постепенно разделяет тело бактерии на две части, в каждой из которых есть молекула ДНК (7).
Бывает (у сенной палочки), две бактерии слипаются, и между ними образуется перемычка (1,2).

По перемычке ДНК из одной бактерии переправляется в другую (3). Оказавшись в одной бактерии, молекулы ДНК сплетаются, слипаются в некоторых местах (4), после чего обмениваются участками (5).
Роль бактерий в природе
Круговорот
Бактерии — важнейшее звено общего круговорота веществ в природе. Растения создают сложные органические вещества из углекислого газа, воды и минеральных солей почвы. Эти вещества возвращаются в почву с отмершими грибами, растениями и трупами животных. Бактерии разлагают сложные вещества на простые, которые снова используют растения.
Бактерии разрушают сложные органические вещества отмерших растений и трупов животных, выделения живых организмов и разные отбросы. Питаясь этими органическими веществами, сапрофитные бактерии гниения превращают их в перегной. Это своеобразные санитары нашей планеты. Таким образом, бактерии активно участвуют в круговороте веществ в природе.
Почвообразование
Поскольку бактерии распространены практически повсеместно и встречаются в огромном количестве, они во многом определяют различные процессы, происходящие в природе. Осенью опадают листья деревьев и кустарников, отмирают надземные побеги трав, опадают старые ветки, время от времени падают стволы старых деревьев. Всё это постепенно превращается в перегной. В 1 см 3 . поверхностного слоя лесной почвы содержатся сотни миллионов сапрофитных почвенных бактерий нескольких видов. Эти бактерии превращают перегной в различные минеральные вещества, которые могут быть поглощены из почвы корнями растений.
Некоторые почвенные бактерии способны поглощать азот из воздуха, используя его в процессах жизнедеятельности. Эти азотофиксирующие бактерии живут самостоятельно или поселяются в корнях бобовых растений. Проникнув в корни бобовых, эти бактерии вызывают разрастание клеток корней и образование на них клубеньков.
Эти бактерии выделяют азотные соединения, которые используют растения. От растений бактерии получают углеводы и минеральные соли. Таким образом, между бобовым растением и клубеньковыми бактериями существует тесная связь, полезная как одному, так и другому организму. Это явление носит название симбиоза.
Благодаря симбиозу с клубеньковыми бактериями бобовые растения обогащают почву азотом, способствуя повышению урожая.
Распространение в природе
Микроорганизмы распространены повсеместно. Исключение составляют лишь кратеры действующих вулканов и небольшие площадки в эпицентрах взорванных атомных бомб. Ни низкие температуры Антарктики, ни кипящие струи гейзеров, ни насыщенные растворы солей в соляных бассейнах, ни сильная инсоляция горных вершин, ни жёсткое облучение атомных реакторов не мешают существованию и развитию микрофлоры. Все живые существа постоянно взаимодействуют с микроорганизмами, являясь часто не только их хранилищами, но и распространителями. Микроорганизмы — аборигены нашей планеты, активно осваивающие самые невероятные природные субстраты.
Микрофлора почвы
Количество бактерий в почве чрезвычайно велико — сотни миллионов и миллиардов особей в 1 грамме. В почве их значительно больше, чем в воде и воздухе. Общее количество бактерий в почвах меняется. Количество бактерий зависит от типа почв, их состояния, глубины расположения слоёв.
На поверхности почвенных частиц микроорганизмы располагаются небольшими микроколониями (по 20-100 клеток в каждой). Часто они развиваются в толщах сгустков органического вещества, на живых и отмирающих корнях растений, в тонких капиллярах и внутри комочков.
Микрофлора почвы очень разнообразна. Здесь встречаются разные физиологические группы бактерий: бактерии гниения, нитрифицирующие, азотфиксирующие, серобактерии и др. среди них есть аэробы и анаэробы, споровые и не споровые формы. Микрофлора — один из факторов образования почв.
Областью развития микроорганизмов в почве является зона, примыкающая к корням живых растений. Её называют ризосферой, а совокупность микроорганизмов, содержащихся в ней, — ризосферной микрофлорой.
Микрофлора водоёмов
Вода — природная среда, где в большом количестве развиваются микроорганизмы. Основная масса их попадает в воду из почвы. Фактор, определяющий количество бактерий в воде, наличие в ней питательных веществ. Наиболее чистыми являются воды артезианских скважин и родниковые. Очень богаты бактериями открытые водоёмы, реки. Наибольшее количество бактерий находится в поверхностных слоях воды, ближе к берегу. При удалении от берега и увеличении глубины количество бактерий уменьшается.
Чистая вода содержит 100-200 бактерий в 1 мл., а загрязнённая — 100-300 тыс. и более. Много бактерий в донном иле, особенно в поверхностном слое, где бактерии образуют плёнку. В этой плёнке много серо- и железобактерий, которые окисляют сероводород до серной кислоты и тем самым предотвращают замор рыбы. В иле больше спороносных форм, в то время как в воде преобладают неспороносные.
По видовому составу микрофлора воды сходна с микрофлорой почвы, но встречаются и специфические формы. Разрушая различные отбросы, попавшие в воду, микроорганизмы постепенно осуществляют так называемое биологическое очищение воды.
Микрофлора воздуха
Микрофлора воздуха менее многочисленна, чем микрофлора почвы и воды. Бактерии поднимаются в воздух с пылью, некоторое время могут находиться там, а затем оседают на поверхность земли и гибнут от недостатка питания или под действием ультрафиолетовых лучей. Количество микроорганизмов в воздухе зависит от географической зоны, местности, времени года, загрязнённостью пылью и др. каждая пылинка является носителем микроорганизмов. Больше всего бактерий в воздухе над промышленными предприятиями. Воздух сельской местности чище. Наиболее чистый воздух над лесами, горами, снежными пространствами. Верхние слои воздуха содержат меньше микробов. В микрофлоре воздуха много пигментированных и спороносных бактерий, которые более устойчивы, чем другие, к ультрафиолетовым лучам.
Микрофлора организма человека
Тело человека, даже полностью здорового, всегда является носителем микрофлоры. При соприкосновении тела человека с воздухом и почвой на одежде и коже оседают разнообразные микроорганизмы, в том числе и патогенные (палочки столбняка, газовой гангрены и др.). Наиболее часто загрязняются открытые части человеческого тела. На руках обнаруживают кишечные палочки, стафилококки. В ротовой полости насчитывают свыше 100 видов микробов. Рот с его температурой, влажностью, питательными остатками — прекрасная среда для развития микроорганизмов.
Желудок имеет кислую реакцию, поэтому основная масса микроорганизмов в нём гибнет. Начиная с тонкого кишечника реакция становится щелочной, т.е. благоприятной для микробов. В толстых кишках микрофлора очень разнообразна. Каждый взрослый человек выделяет ежедневно с экскрементами около 18 млрд. бактерий, т.е. больше особей, чем людей на земном шаре.
Внутренние органы, не соединяющиеся с внешней средой (мозг, сердце, печень, мочевой пузырь и др.), обычно свободны от микробов. В эти органы микробы попадают только во время болезни.
Бактерии в круговороте веществ
Микроорганизмы вообще и бактерии в частности играют большую роль в биологически важных круговоротах веществ на Земле, осуществляя химические превращения, совершенно недоступные ни растениям, ни животным. Различные этапы круговорота элементов осуществляются организмами разного типа. Существование каждой отдельной группы организмов зависит от химического превращения элементов, осуществляемого другими группами.
Круговорот азота
Циклическое превращение азотистых соединений играет первостепенную роль в снабжении необходимыми формами азота различных по пищевым потребностям организмов биосферы. Свыше 90% общей фиксации азота обусловлено метаболической активностью определённых бактерий.
Круговорот углерода
Биологическое превращение органического углерода в углекислый газ, сопровождающееся восстановлением молекулярного кислорода, требует совместной метаболической активности разнообразных микроорганизмов. Многие аэробные бактерии осуществляют полное окисление органических веществ. В аэробных условиях органические соединения первоначально расщепляются путём сбраживания, а органические конечные продукты брожения окисляются далее в результате анаэробного дыхания, если имеются неорганические акцепторы водорода (нитрат, сульфат или СО 2).
Круговорот серы
Для живых организмов сера доступна в основном в форме растворимых сульфатов или восстановленных органических соединений серы.
Круговорот железа
В некоторых водоёмах с пресной водой содержатся в высоких концентрациях восстановленные соли железа. В таких местах развивается специфическая бактериальная микрофлора — железобактерии, окисляющие восстановленное железо. Они участвуют в образовании болотных железных руд и водных источников, богатых солями железа.
Бактерии являются самыми древними организмами, появившимися около 3,5 млрд. лет назад в архее. Около 2,5 млрд. лет они доминировали на Земле, формируя биосферу, участвовали в образовании кислородной атмосферы.
Бактерии являются одними из наиболее просто устроенных живых организмов (кроме вирусов). Полагают, что они - первые организмы, появившиеся на Земле.
Оглавление темы "Систематика живых организмов. Таксономия. Номенклатура. Классификация микроорганизмов.":1. Систематика живых организмов. Таксономия. Номенклатура.
2. Классификация микроорганизмов. Принципы классификации микроорганизмов. Систематика микроорганизмов. Естественная (филогенетическая) систематика микроорганизмов.
3. Искусственная (ключевая) систематика микроорганизмов. Определитель бактерий Берджи.
4. Принципы таксономии микроорганизмов. Принципы номенклатуры микроорганизмов. Категории таксономической иерархии. Названия таксонов у микроорганизмов.
5. Систематика вирусов. Особенности классификации вирусов. Основные критерии таксономической классификации вирусов.
6. Систематика бактерий. Окраска по Граму. Грамположительные бактерии. Грамотрицательные бактерии. Кислотоустойчивые бактерии.
8. Определитель Берджи. Группы бактерий определителя Берджи.
9. Систематика грибов. Принципы классификации грибов. Зигомицеты. Аскомицеты. Базидиомицеты. Дейтеромицеты. Кодекс названий грибов.
10. Систематика простейших. Принципы классификации простейших. Тип Sarcomastigophora. Тип Ciliophora. Тип Apicomplexa.
Важным дифференцирующим признаком является подвижность. В соответствии со способом передвижения выделяют скользящие бактерии , перемещающиеся за счёт волнообразных сокращений тела, и плавающие бактерии , движение которых обеспечивают жгутики или реснички.
Способность к спорообразованию бактерий
Для классификации некоторых бактерий учитывают их способность к спорообразованию, размеры спор и их расположение в клетке.
Физиологическая активность бактерий
Физиологическая активность является не менее важным отличительным признаком. Бактерии разделяют по способу питания, по типу получения энергии (дыхание, брожение, фотосинтез), по отношению к рН с указанием пределов устойчивости и оптимума роста и т.д. Наиболее важным критерием считают отношение к кислороду.
Аэробные бактери и используют молекулярный 02 в качестве конечного акцептора электронов в процессе дыхания. Большинство бактерий обладают связанной с мембраной цитохром-С-оксидазой, играющей ведущую роль в электронотранспортной цепи. Для выявления фермента применяют оксидазный тест, основанный на способности бесцветного вещества N,N-диметил-р-фенилендиамина приобретать при восстановлении малиновый цвет.
Анаэробные бактерии не утилизируют молекулярный 02 в качестве конечного акцептора электронов. Подобные бактерии получают энергию либо в процессе брожения, где конечными акцепторами электронов служат органические соединения, либо при анаэробном дыхании, используя отличный от кислорода акцептор электронов (например, NO", SO4- или Fe3+). Факультативные бактерии могут получать энергию либо в процессе дыхания, либо при брожении в зависимости от наличия или отсутствия кислорода в окружающей среде.
Биохимические свойства бактерий
Для дифференцировки бактерий изучают их способность ферментировать углеводы, образовывать различные продукты (сероводород, индол) или гидролизовать белки.
Антигенные свойства бактерий
Антигенные свойства бактерий различных бактерий специфичны и связаны с особенностями строения клеточных структур, распознаваемых специальными антисыворотками как антигенные детерминанты. Типирование бактерий по антигенной структуре проводят в реакции агглютинации (РА), смешивая каплю антисыворотки с каплей бактериальной суспензии. При положительной реакции появляются отдельные агрегированные комочки в исходно однородной бактериальной суспензии. Выделяют следующие типы Аг:
родоспецифичные
, выявляемые у всех представителей конкретного рода, включая отдельные штаммы;
видоспецифичные
, выявляемые у отдельных видов и штаммов микроорганизмов;
сероваро- (штаммо-) специфичные, выявляемые у представителей различных подгрупп (штаммов) внутри конкретного вида.
Чувствительность бактерий к бактериофагам
Поскольку вирусы бактерий взаимодействуют только с чувствительными бактериями, то в ряде случаев для дифференцировки бактерий применяют их чувствительность к бактериофагам.

Химический состав бактерий
Важный классификационный признак - суммарный химический состав бактериальных клеток . Обычно определяют содержание и состав Сахаров, липидов и аминокислот в клеточных стенках.
Генетическое родство бактерий
Для филогенетической классификации бактерий лучший и наиболее информативный показатель - генетическое родство. При систематизации бактерий на основании генетического родства учитывают ряд показателей.
Способность обмениваться генетической информацией (например, в процессе трансформации или конъюгации), возможной только между организмами одного рода или вида.
Состав оснований ДНК (отношение гуанин-цитозин: аденин-тимин).
Сходство нуклеиновых кислот, выявляемое методом гибридизации.
"Золотой стандарт
" в определении степени родства среди бактерий - установление сходства в последовательности нуклеиновых кислот. Однако в результате обмена генетической информацией генотип бактерий может существенно изменяться, поэтому подобный подход к систематике оказывается на практике неудобным.